
Полная версия:
Garden Birds
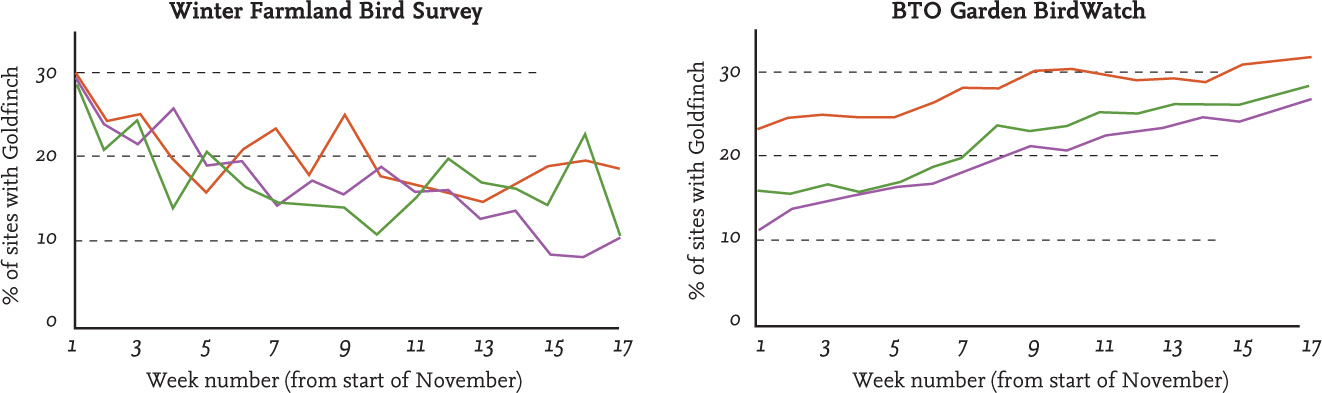
FIG 32. Data from two BTO surveys suggest a movement into gardens by Goldfinches as the winter progresses, something that could be studied in greater detail through the use of ringing or tagging technologies. Data reproduced from BTO with permission.
IS THE PROVISION OF SUPPLEMENTARY FOOD FOR WILD BIRDS A GOOD THING?
Little work has been done to examine differences in the potential costs or benefits of different types of food – at its simplest, high-quality or low-quality foods. We are not yet in a position to be able to determine whether it is better to encourage people to feed wild birds, regardless of the type of food that they provide, because it fosters a sense of connection with the natural world, or discourage the provision of feeding altogether if we cannot provide foods that are entirely suitable for the species that we are trying to help. We provide supplementary food and garden for wildlife more generally because we see these activities as being beneficial for wildlife. However, while these resource subsidies and habitat modifications may provide opportunities for birds and other wildlife within our increasingly urbanised landscapes, they may also fundamentally alter ecological processes and community structure (Shochat, 2004). Understanding these potential impacts requires new research.
Later in this chapter we will look in more detail at some of the costs and benefits for birds of feeding on food provided at garden feeding stations (or experimentally in other habitats). One study, however, is worth mentioning here since it nicely summarises some of these costs and benefits. The study in question was carried out over three years and examined how bird feeding impacts bird health – something that involved looking at body condition, stress, antioxidant levels, nutritional condition, immune function and levels of disease – for wild bird communities in two forested sites, one where food was provisioned and one where it wasn’t (Wilcoxen et al., 2015). Wilcoxen’s study revealed some clear and beneficial effects; birds at the fed site had significantly larger fat stores, greater antioxidant capacity and reduced stress. In addition, fed birds also showed better body condition and better nutritional condition, though not in every year. However, fed birds were more likely to be suffering from disease. Ten months after the feeding experiment had ended, the researchers repeated the sampling carried out at the two sites; this time the differences had gone, adding further supporting evidence that the differences had been due to the provision of food. Feeding does, it seems, deliver some health benefits, but these come with a certain level of cost.
DOES PROVIDING FOOD ENCOURAGE DEPENDENCY?
One of the most common criticisms of the practice of providing food for wild birds is that it causes dependency, prompting questions as to what happens to birds if feeding stops, and are individuals that use feeders regularly able to then recognise natural foods or have the skills needed to exploit them? Little work has been done on the question of dependency, in part a reflection of the often quoted work done by Brittingham & Temple (1992b) – see below – and in part a reflection of knowledge gained on how small birds track different feeding opportunities.
Brittingham and Temple recognised that two different types of dependency may occur. Over the short term, birds visiting a feeder may come to expect that particular location to provide food; if the feeder is left empty or is removed, then the bird may suffer. Over the longer term, those individuals that are heavy users of feeders may lose the ability to recognise naturally occurring food items, lose the skills to handle them and suffer from reduced survival rates. Working on two resident populations of Black-capped Chickadee, Brittingham and Temple tested whether the population that had utilised feeders in the past experienced lower survival rates than a population that had never been exposed to them during a winter when feeders were not available to either population. The researchers found no difference between the average monthly survival rates of the two populations and concluded that there was no evidence for dependency in this species.

FIG 33. The question of whether wild birds become dependent on garden feeding stations has been largely answered by scientific studies; these indicate that small birds are aware of other feeding opportunities and will quickly turn to these if food is no longer available at a given site. (John Harding)
The question of whether the sudden cessation of food provision can have a negative effect on individual birds has not been tested but is not thought to be a particular issue. The thinking that supports this view comes from the work on chickadees, which demonstrates that individual birds track a number of food patches at the same time, sampling these no matter how abundant food is at any one patch (Brittingham & Temple, 1992b). For a chickadee living in the northern part of North America – and being one of the smallest bird species to remain so far north during the winter months – tracking a range of food patches has clear survival benefits; if conditions change and one patch is no longer available, individuals already know where other patches are located. The sudden loss of a feeding opportunity might be more of an issue for other species, but one would expect the urban environment and its gardens to provide multiple feeding opportunities within a short distance of each other. This is something that has been examined, to a small degree, through the work of Patrick Thompson, who looked at the effect of interrupting regular food provision in gardens on the numbers and behaviour of House Sparrows and Greenfinches (Thompson, 1989). Thompson’s work revealed that the daily pattern of feeding station use was unaffected by the number of neighbouring gardens providing food. However, the numbers of individuals using the experimental garden feeding station was dependent on food provision in neighbouring gardens. When the number of neighbouring gardens providing food was reduced, there was an increase in activity within the experimental garden, where feeding had continued.
THE VALUE OF GARDEN FEEDING STATIONS FOR WILD BIRDS
Many of the individuals providing food for birds at their garden feeding stations during the winter months do so because they believe that it will help the birds through a difficult period of the year. The provision of food might improve the survival rates of fed individuals, but to what degree. Work on Black-capped Chickadees in North America has revealed that supplementary feeding can make a significant difference to overwinter survival. The fed chickadees studied by Brittingham & Temple (1988a) were significantly heavier than unfed birds and, while this difference in weight was very small (equivalent to just 1 per cent of body weight and representing two hours of foraging time), it made a big difference to their longer-term survival rates. Fed individuals had a 70 per cent survival rate through to the end of the winter and beyond, compared with just 30 per cent in unfed birds. Work on Willow Tits and Crested Tits, carried out in coniferous woodland near Gothenburg in Sweden by Jansson et al. (1981), also found that the provision of additional food resource improved winter survival significantly. In this case, the three researchers noted that the provisioned food helped to reduce predation risk by allowing individuals to spend more time in far more protected places than they would frequent under natural food conditions.
FOOD PROVISION AND MEASURES OF REPRODUCTIVE SUCCESS
Various studies have demonstrated that increasing the availability of food resources through the provision of supplementary food can advance the onset of breeding, prolong the period over which breeding takes place and increase both the number of young raised per breeding attempt and the number of breeding attempts made. Work on Florida Scrub-jays Aphelocoma coerulescens (Fleischer et al., 2003; Schoech et al., 2004) has, for example, revealed that populations with access to supplementary food can start breeding up to 17 days earlier than populations without such access. Similar findings have been revealed for Australian Magpie (Rollinson & Jones, 2002).
The timing of breeding
The work on Florida Scrub-jays reveals both the differences that supplementary food provision can make to the timing of breeding and some of the mechanisms behind it. Suburban scrub-jay populations have access to supplemented food in the form of peanuts, cracked corn, birdseed, bread and pet food, with up to 30 per cent of their diet made up of such items (Fleischer et al., 2003). This food forms a predictable resource, which is in contrast to the unpredictable nature of resources experienced by wildland populations. Predictability appears to be important and probably explains why Fleischer et al. (2003) found their suburban scrub-jay populations to be consistent in the timing of their breeding across years, while wildland populations varied. Having a predictable food source in the form of supplemented food may well explain why suburban scrub-jay populations spend less time foraging and more time perching than wildland birds – scrub-jays use a sentinel system, with perching birds alerting other individuals to the presence of potential predators. Suburban scrub-jays also handled more food items per hour, suggesting that their foraging was more efficient. Interestingly, while food consumption rates were not found to vary between the two habitats, those individuals that were the most efficient foragers were also the earliest breeders.
So what does the work on scrub-jays tell us about the effects of supplementary feeding on the timing of breeding? If, as the work of Fleischer et al. (2003) suggests, foraging efficiency increases through access to garden feeding stations, the predictability of this significant food resource may provide the birds with a suitable cue for the appropriate timing of breeding. Of course, the supplementary food might influence the timing of breeding in other ways, perhaps by altering female body condition or behaviour. These two possibilities are known as the ‘anticipation hypothesis’, where females use food availability as a cue for the onset of breeding, and the ‘constraint hypothesis’ – where food availability constrains the female’s ability to produce eggs.
Earlier work by the same research group (Schoech & Bowman, 2001) revealed few differences in female body condition prior to breeding between suburban and wildland populations, suggesting instead that the rate of food intake or time spent foraging – both of which are shaped by food predictability – were more important. Later work on Florida Scrub-jays has attempted to look in greater detail at some of the underlying mechanisms, focussing on protein in the food taken by the birds and also monitoring the levels of key reproductive hormones. Protein is thought to be of particular importance, with evidence suggesting that wildland scrub-jay populations are protein limited – they have lower levels of plasma protein than suburban birds (Schoech & Bowman, 2003). Wildland scrub-jays feed on acorns, the tannic acids in which have been shown to reduce the ability to assimilate proteins. Schoech & Bowman provided their study populations with supplementary food of two different types; one was high in fat and high in protein (‘HFHP’), and one was high in fat but low in protein (‘HFLP’). Scrub-jays in both treatment groups bred earlier than those in an unfed control group, but birds in the HFLP group bred later than those in the HFHP group.
The researchers then looked at testosterone levels in male scrub-jays, estradiol levels in females, and corticosterone levels in both sexes. The HFHP males had higher testosterone levels than either the HFLP or control group males, but no difference was found in estradiol levels in the females. Estradiol mediates maturation of the ovarian follicles and also appears to stimulate nest building and solicitation behaviour, so the researchers had hypothesised that there would be differences in this key hormone between groups. That the high-protein diet influenced a key male hormone rather than a key female hormone could suggest that male scrub-jays play a larger role in determining when reproduction commences than the females. The corticosterone findings were particularly interesting because they revealed that the levels of this ‘stress’-related hormone were lower in suburban populations of the scrub-jays than in either the HFHP, HFHP or control group – though HFHP birds had lower corticosterone levels than HFLP or control group birds. This suggests that the predictability of food resources may influence corticosterone levels and that corticosterone may play a role in the timing of reproduction. This work highlights the complex pathways by which something as apparently simple as the provision of additional food resources may have on the timing of breeding in birds.
Work on species found breeding in UK gardens has generally identified a similar pattern, with the provision of supplementary food leading to earlier laying in Blue Tits in Northern Ireland (Robb et al., 2008a). Robb’s Blue Tits laid an average of 2.5 days earlier than unfed birds. However, Plummer et al. (2013), working on Blue Tits, found that laying dates did not differ significantly between their fed and unfed groups. It is worth just noting that most of these studies have been carried out in woodland habitats rather than in gardens, something that reflects the very great difficulty in running controlled experiments within an urban setting (see Chapter 6). Elsewhere, the laying dates of Willow Tit and Crested Tit were advanced by 2–5 and 5–8 days respectively when provided with sunflower seed and a tallow-based mix containing soy protein, wheat germ, sunflower seed, vitamins and minerals (Brömssen & Jansson, 1980). In a review of 59 studies, Robb et al. (2008b) found that 58 per cent reported significantly earlier laying dates in supplemented populations. Some studies, but not others, suggest that supplementary food may shorten incubation periods (Nilsson & Smith, 1988; Nager et al., 1997, Harrison et al., 2010). The Harrison et al. (2010) study revealed that supplemented Great Tits reduced their incubation period in each of the three study years, while Blue Tits did so in two of the three years.

FIG 34. The presence of supplementary food at garden feeding stations can advance the timing of breeding in species like Blue Tit, something that has been proved experimentally here in the UK. (Jill Pakenham)
Clutch size, egg and offspring quality and fledging success
The formation of an egg is not only costly in terms of energy but also in terms of nutrients, such as protein. If females are limited by resources as they go into the egg laying period, then it follows that the provision of suitable resources may lead to an increase in clutch size. However, the evidence for this is mixed, with some authors finding an effect (e.g. Soler & Soler, 1996) but not others (Brömssen & Jansson, 1980; Svensson & Nilsson, 1995). In one case, the provision of supplementary food appeared to reduce clutch size significantly in Blue Tits and Great Tits breeding in a Worcestershire woodland (Harrison et al., 2010). The Harrison study revealed an interesting mix of results; in addition to the significant reduction in clutch size and the shortened incubation periods noted above, food supplementation from several weeks pre-laying through to hatching appeared to reduce hatching success in Blue Tits, but not Great Tits. Robb et al.’s review, mentioned earlier, found that in 44 studies that dealt with food supplementation and both laying dates and clutch size, 12 reported increases in both and 16 reported increases in clutch size alone (Robb et al., 2008b).

FIG 35. Supplementary food has been shown to influence the number and quality of offspring produced by birds like Great Tit. (Mike Toms)
The provision of supplementary food, just prior to and during egg laying, has been shown to result in increased deposition of carotenoids within the yolk of an egg, leading to increased hatching success (Møller et al., 2008) and, subsequently, higher rates of adult survival (McGraw et al., 2005). Chicks hatched from eggs with high levels of antioxidants have been shown to have lower rates of mortality in the days following hatching (Royle et al., 2011), so micronutrients stored by the female bird and derived from supplementary food, could deliver benefits to the resulting chicks. However, the findings of various studies show some differences. Supplementary feeding during the winter months – this time with fat – has been shown to affect investment in egg production, resulting in a smaller relative yolk mass in larger eggs and reduced yolk carotenoid concentration in early breeding Blue Tits (Plummer et al., 2013); these negative effects were absent in Blue Tits fed with fat plus vitamin E (see below). Egg size increases were reported in 38 per cent of the studies examined by Robb et al. (2008b). Returning to Florida Scrub-jays, Jim Reynolds (Reynolds et al., 2003) found that females fed with a high-fat high-protein diet produced heavier third eggs, containing more water and more protein than the eggs of unsupplemented individuals.
Of course, one of the key figures when discussing breeding success is the number of chicks that fledge from the nest; while this is obviously influenced by some of the factors we have just discussed – such as the number of eggs – it is worth looking at whether supplementary feeding influences this figure. Working on a Blue Tit population in Northern Ireland, Gillian Robb and colleagues found that supplementary feeding significantly affected the numbers of chicks that fledged, such that individuals for which peanuts had been provided during the winter months fledged almost one extra chick per nest compared to those without access to the food (Robb et al., 2008a). Although Brömsson & Jansson (1980) did not find any effects of food supplementation on clutch size or the number of fledglings in Willow Tit or Crested Tit, they did find that Crested Tit nestlings were significantly heavier within the fed study site and that there was a tendency to produce second clutches more often in the experimental area.
One area where concern has been raised about the practice of feeding garden birds during the breeding season is in relation to nestling health and well-being. It has been suggested that garden birds might feed their nestlings on artificial foods provided at garden feeding stations in preference to the natural, predominantly invertebrate-based, foods that they actually need. This suggests that garden birds are either lazy or unable to identify foods as being unsuitable for their nestlings. However, given what we know about food selection behaviour in wild birds, and which has been presented elsewhere in this chapter, both of these suggestions seem unlikely. Cowie & Hinsley (1988a; 1988b) examined nestling diet in a population of suburban Blue Tits and Great Tits, using nest cameras to identify the food provided by parent birds and how this was delivered. The cameras revealed that, on average, 15 per cent of the nestling diet of each of the two species was made up of artificial foods, notably bread, fat and peanuts. The amount provided varied between pairs, ranging from 6 per cent to 34 per cent in Great Tit and from 6 to 28 per cent in Blue Tit. Despite this, the researchers failed to find any relationship between the amount of artificial food provided and measures of nestling health. That adult garden birds are able to make appropriate decisions about the food they provide to their chicks can be seen from observations that if additional insect food is made available to nesting tits, or if their brood size is artificially reduced by removing some of the nestlings, the parent birds respond by dropping artificial foodstuffs from their diet and increasing the numbers of invertebrates provisioned.

FIG 36. Adult birds appear to be able to identify appropriate foods for their nestlings and fledglings, with most species feeding their young on invertebrate prey while they are still in the nest, and making use of bird table fare as the young develop towards independence. (John Harding)
Supplementary food, such as that presented to birds within the garden environment, may have consequences for other measures of breeding success, and indeed for other aspects of the breeding cycle. Studies have revealed that nesting adults, provided with additional food, will often reduce foraging effort, something that can provide time for other activities – such as predator vigilance (Arcese & Smith, 1988). It can also have some rather surprising effects. Conservationists, working to secure a future for the Kakapo Strigops habroptilus through supplementary feeding of the worryingly small female population, discovered to their horror that the quality of food being provided led to a male-biased sex ratio in the resulting chicks. It is thought that this came about because the female is the heterogametic sex – it has two different sex chromosomes – and can in some instances determine the sex ratio of offspring at the point of fertilisation. A change to the food quality being provided saw a more balanced sex ratio restored (Robertson et al., 2006).
Diet and nutrition have also been linked to important traits associated with reproduction, including mate selection. Mate selection is often based around traits that indicate the quality of a potential mate. Such traits, which include plumage colouration and song complexity, are often nutritionally or energetically costly to maintain.
THE HEALTH EFFECTS OF PROVIDING FOOD
Until recently, very little work had been done on the nutritional composition of the foods being provided at garden feeding stations; even now we are only just beginning to explore the health and other impacts of such provisioning. We know that the proteins and lipids found in naturally occurring foods play an important role in the control of reproduction, underlining that we need to establish how supplemented foods differ from what birds would be eating normally and what, if any, effects such differences have on the health and behaviour of individual birds. We also know that the provision of supplementary food can support birds as they replace lost feathers, something tested experimentally by Grubb & Cimprich (1990).

FIG 37. Although the black bib of male House Sparrows has an important function, signalling status, it is not as sensitive to nutritional conditions as one might expect. (John Harding)
A poor diet can have implications for the health of a wild bird, perhaps increasing susceptibility to disease or resulting in poor-quality plumage or other abnormalities. In geese and birds of prey, nutritional imbalance has been suggested as the primary cause of ‘angel wing’ disorder, while the effects of malnutrition on plumage characteristics have been seen in Brown-headed Cowbirds Molothrus ater (McGraw et al., 2002). Interestingly, McGraw’s work, which looked at both Brown-headed Cowbird and House Sparrow, found that important signalling plumage – the melanin-based black bib of male House Sparrows and the brown hood of the cowbirds – was not affected by nutritional stress. That such melanin-based ornaments are less sensitive to nutritional conditions during moult suggests these may signal hormonal status and/or competitive ability rather than represent an accurate signal of health.
The provision of meat at garden feeding stations, which is widely practised in Australia but not the UK, could have implications for the health of those birds feeding upon it. It has been demonstrated that birds like Australian Magpie, taking meat from garden feeding stations, have elevated levels of plasma cholesterol and fatty acids (Ishigame et al., 2006). A secondary concern with meat is that it has an unbalanced calcium to phosphorous ratio – calcium absorption can be hindered by high phosphorus levels – which can lead to a range of skeletal or physiological problems.

