
Полная версия:
Garden Birds
The reliance on foods provided at garden feeding stations may lead to a dietary imbalance, directly affecting an individual bird, and/or impairing its reproductive potential during a subsequent breeding season. Where events in one season influence an individual’s performance in a later season, this is known as a ‘carry-over effect’. Such effects have received increasing attention over the last few years, particularly in relation to supplementary feeding. Another potential effect of food provision at garden feeding stations is that it may alter population structure, perhaps by enabling low quality individuals to recruit into the breeding population. The phenotypic quality of individuals is strongly influenced by oxidative stress, with this oxidative stress reflecting the imbalance between the harmful reactive oxygen species (known as ROSs) produced as by-products of metabolism and the body’s antioxidant defence system (Selman et al., 2012).
Carry-over effects are often talked about in terms of energetics, with macronutrients – such as fats – thought to be of particular importance. However, small birds, such as those visiting garden feeding stations in the UK, lack the capacity to store large amounts of fat, meaning they have to rely on daily food intake to meet the energetic demands imposed by reproduction. Because of this, it might be that micronutrients – such as antioxidants – are more important; these may be stored in the liver and any subcutaneous fat to deliver reserves that can be utilised during a subsequent breeding season or at other times when they are needed (Metzger & Bairlein, 2011).

FIG 38. Adult Great Spotted Woodpeckers bring their chicks to garden feeding stations soon after they have left the nest. Presumably knowledge of a reliable food source will help the young woodpeckers as they become independent. (Jill Pakenham)
Work by Kate Plummer and colleagues has sought to establish the extent to which supplementary feeding produces the individual- and population-level effects we have just described (Plummer et al., 2018). Plummer used different woodland populations of Blue Tits to compare the effects of providing fat, and fat plus vitamin E (an antioxidant), with a control population of unfed birds. Feeding was only carried out during the winter months, ending at least a month before the tits began egg laying. Provisioning with fat and vitamin E improved the survival, recruitment and breeding condition of birds that had been in significantly poorer condition prior to feeding; provisioning with fat only was found to have a detrimental impact on breeding birds. Because birds that had been supplied with fat and vitamin E were found to have lower levels of carotenoids in their breast feathers – lower levels of carotenoids being indicative of lower quality individuals – Plummer was able to conclude that supplementing with vitamin E and fat in winter had altered the survival and recruitment prospects of these lower quality individuals. It appears, therefore, that the provision of supplementary foods during the winter months can alter both the structure of the breeding population and the condition of individual breeding birds. Such effects can also have consequences beyond the breeding season where, for example, food provision alters levels of oxidative damage; in Plummer’s study, individuals with higher blood plasma concentrations of malondialdehyde (indicative of oxidative damage) produced offspring that were structurally smaller and suffered from reduced fledging success.
The importance of antioxidants, like vitamin E, can also be seen from Plummer’s work on yolk mass, mentioned earlier. While Plummer et al. (2013) found that winter provisioning with fat subsequently impaired an individual’s ability to acquire, assimilate and/or mobilise key resources for yolk formation, this was not the case where vitamin E was also included in the food presented. A high-fat diet, such as that potentially obtained from the food provided at garden feeding stations, may well increase the requirement for antioxidants in order to combat the greater levels of oxidative damage associated with a diet rich in fats.
FOOD PROVISION, POPULATION SIZE AND COMMUNITY STRUCTURE
Plummer’s work shows that the effects of food provision can potentially pass from one generation to the next, and alter the composition of breeding populations in subsequent seasons, but what about its impact at a wider community level? Work carried out in the Netherlands has revealed how the provision of supplementary food can lead to an increase in local population size, and how the scale of the effect can change in relation to the availability of wider food resources (van Balen, 1980). Research by van Balen monitored two populations of Great Tits, breeding in similar woodlands just 7 km apart; before introducing a regime of winter feeding, the two populations were similar and had been fluctuating more or less in parallel over several years. Following the introduction of feeding, the number of breeding pairs in the fed woodland increased to become 40 per cent higher than that in the unfed woodland. Not only this, but the scale of the difference varied according to the size of the beechmast crop; in poor beechmast years, the fed population was almost double that of the unfed population the following breeding season; following years with a good mast crop, there was little difference between the numbers of Great Tits breeding in the two woods. A similar finding was noted by Källander (1981), working in Sweden, whose first year of study (1969–70) coincided with a particularly severe winter. Despite this, Great Tits in a wood provisioned with sunflower seeds increased in number, while those in an unfed wood declined. The following year, in which there was a good crop of beechmast, saw populations in both woods increase.
If the provision of food at garden feeding stations leads to an increase in the numbers of birds locally, then we might expect to see a relationship between feeder density and bird numbers across individual towns and cities. There is some evidence of this from the work carried out in Sheffield (Fuller et al., 2008; 2012), with the density of three of the seven urban-adapted species studied shown to be positively related to the density of bird feeders. House Sparrow showed the strongest pattern, with 57 per cent of the variation in House Sparrow densities explained by the density of bird feeders; the other two species were Blackbird and Starling. Interestingly, given the frequency with which Blue Tits and Great Tits are seen at garden feeding stations, it was surprising that they did not show such a relationship, yet Blackbird did.
Some of the strongest evidence for community-level effects comes from work carried out in New Zealand, where most of the native bird species able to persist within urban areas are either predominantly insectivorous (e.g. Grey Warbler Geryone cigata), frugivorous (e.g. New Zealand Pigeon Hemiphaga novaeseelandiae) or nectarivorous (e.g. Tūī Prosthemadera novaeseelandiae). In contrast, many of the introduced species are either granviorous (e.g. House Sparrow and Spotted Dove) or adaptable enough to take bird table fare. Much of the food provided at garden feeding stations in New Zealand would appear to favour the introduced species, something confirmed by Galbraith et al. (2015) through a feeding experiment. The introduction of feeding altered the community structure, with significant increases seen in the abundance of House Sparrow and Spotted Dove, and to a lesser degree in Starling and Song Thrush. The introduction of feeding was also shown to have a negative effect on the abundance of the native Grey Warbler. Once the feeding experiment ended, virtually all of the observed changes in community structure disappeared. Observational work on these birds (Galbraith et al., 2017b) revealed that House Sparrow and Spotted Dove dominated the feeding stations.
Supplementary feeding in the Sydney area, Australia, together with the establishment of both native and non-native fruiting shrubs in gardens, is thought to have been a significant factor in the major changes seen over the past 100 years in the diversity and abundance of parrots (Burgin & Saunders, 2007). The provision of food at garden feeding stations is thought to have aided colonisation and range expansion of the Collared Dove within the UK. Within North America, the northwards range expansions of House Finch, Tufted Titmouse Baeolophus bicolor and Northern Cardinal have also been attributed to the food available at garden feeding stations during the winter months.

FIG 39. The presence of garden feeding stations is thought to have aided the successful colonisation of the UK by Collared Doves. (John Harding)
INFLUENCE OF SUPPLEMENTARY FOOD ON NEST PREDATION
We have already seen how the provision of supplementary food can alter the timing of breeding and community structure. A special case of the latter relates to the ways in which the provision of supplementary foods can influence relationships between predators and their prey. This is of particular relevance when discussing urban and suburban gardens because anthropogenic foods have become a defining characteristic of urbanised landscapes (Warren et al., 2006). The nature of interactions between breeding birds and their predators may be especially influenced by the provision of such supplementary foods because both songbirds and their generalist predators readily exploit these resources, something that can lead to them occupying the same urban sites. BTO work, in the form of a meta-analysis led by Dan Chamberlain, failed to reveal any strong underlying pattern to the effects of urbanisation – and the associated anthropogenic food sources – on the relationship between predators and levels of nest predation (Chamberlain et al., 2009a), but there have been some useful field experiments on the subject.

FIG 40. There is little evidence that the presence of bird feeders and their use by Grey Squirrels leads to increased levels of nest predation in the local area, but such an effect has been seen for other predators. (John Harding)
Supplementary food may reduce levels of nest predation by providing an alternative food supply for would-be nest predators; alternatively, it may increase levels of nest predation by attracting predators or elevating their population densities. Jennifer Malpass and her colleagues investigated how the presence of bird feeders affected predator abundance and nest survival of American Robin Turdus migratorius and Northern Cardinal across seven neighbourhoods in Ohio, US (Malpass et al., 2017). Malpass found that the relative abundance of both Brown-headed Cowbird and American Crow Corvus brachyrhynchos – known nest predators of the two study species – was positively associated with the number of bird feeders present within a neighbourhood. No similar relationship was found for the other nest predators studied, which included Grey Squirrel Sciurus carolinensis, Blue Jay Cyanocitta cristata and Common Grackle Quiscalus quiscula. While the relative abundance of American Crow and Brown-headed Cowbird was greater in areas with bird feeders, there was no consistent relationship between the numbers of bird feeders and predation of the songbird nests.
DISEASE AND FOOD PROVISION
The presence of supplementary food at garden feeding stations may contribute positively to bird populations by increasing the availability of food during those periods when natural food supplies are in short supply. However, as will be explored in Chapter 4, the provision of food at bird tables and in hanging feeders may also facilitate the spread of disease. The relationships between food provision and disease occurrence can be more complex than they first appear. It is not simply that garden feeding stations attract large numbers of birds, and through this lead to increased opportunities for disease transmission; in some instances the presence of the food can act to ameliorate the impacts of disease. This is something that has been studied in North America, in relation to the outbreak of the bacterial disease mycoplasmal conjunctivitis in North American House Finches (Fischer & Miller, 2015).
Mycoplasmal conjunctivitis impairs vision, making it difficult for affected individuals to find and locate food. If such individuals have access to abundant and predictable food resources, this may afford them with the time and energy needed to recover. Interestingly, there is also some limited evidence that individuals infected at feeding stations may develop less severe symptoms than those infected elsewhere, something that may lead to a faster recovery. Fischer & Miller (2015) used information from the FHWAR (Fishing, Hunting and Wildlife-associated Recreation) census to examine how food provision influenced House Finch populations before and after the emergence of mycoplasmal conjunctivitis. The two researchers found that the availability of bird food appeared to have a positive effect on House Finch populations, with House Finch density prior to the disease outbreak strongly related to the density of US citizens providing food for birds. After the disease arrived, reducing House Finch numbers substantially, the relationship between feeder density and finch numbers remained.
A comparison was then made between pre- and post-disease emergence populations, the researchers seeking to determine whether finch populations declined more strongly where feeder densities were high – which is what you might expect if high densities of feeders lead to high rates of disease transmission and, through this, to a correspondingly greater level of population decline. The results of the study revealed a positive relationship between changes in finch densities and changes in feeder densities between the two periods, indicating that higher densities of feeders actually resulted in lower rates of population decline. This adds weight to the work of Dhondt et al. (2007), suggesting that food provision might act as a ‘crutch’ for sick birds and improve their chances of survival and recovery. Other factors also need to be considered, not least that House Finches may spread mycoplasmal conjunctivitis to other species, including House Sparrow and American Goldfinch.
FOOD PROVISION AND BEHAVIOUR
As we have already discovered, food resources are central to many different aspects of a bird’s life, so it is little wonder then that the provision of food can alter behaviour. The area where we might expect to see clear evidence for changes in behaviour is in relation to territory, since territory provides a means of partitioning and defending resources, including food. Black-capped Chickadees show a degree of territoriality during the winter months, defending small foraging patches from intruders. However, as Wilson (2001) discovered, such territoriality breaks down when a significant food source is added to a foraging patch, the resident flock (typically the resident mated pair and six to ten first winter birds) unable to defend the new resource against the large numbers of other chickadees wanting to make use of it.

FIG 41. When supplementary food is available, both Blue Tits and Great Tits are less likely to join the mobile foraging flocks that are a feature of the early winter in woodland habitats in the UK. (Jill Pakenham)
A feature of the tit species that are familiar to garden feeding stations here in the UK is their willingness to form mixed-species flocks during the autumn and winter months, a behaviour that also occurs in other tit species elsewhere in the world. This behaviour is thought to improve an individual’s chances of finding food during those periods when food is scarce and encounter rates low. You might, therefore, expect to see the provision of supplementary food exert an influence on the tendency to form or join mixed-species flocks. This is exactly what Thomas Grubb found, working on tits in deciduous woodland in Oxfordshire (Grubb, 1987). Grubb was interested in whether mixed-species flocking was driven by predation risk, by food availability or by both. His results suggested that food was the primary driver; Blue Tits and Great Tits without access to supplementary food flocked with other species while foraging more often than was the case when supplementary food was available. Interestingly, Long-tailed Tits – which ignored the artificial food – foraged in mixed-species flocks regardless of whether or not provisioned food was available. Work in Japan on Varied Tit Sittiparus varius underlines the behavioural flexibility in whether or not individuals join mixed-species flocks (Kubuta & Nakamura, 2000), revealing that individuals participate in mixed-species flocks to obtain the short-term benefits of increased foraging efficiency but, independent of food provision, they also obtain long-term benefits from the stability of their pair bonds and strong site fidelity.
The studies just mentioned are relevant to the discussion of how the winter provision of food at garden feeding stations may alter the behaviour of the birds that visit. However, garden feeding may also change behaviour during the breeding season, something that may be of particular importance. As we shall see in our examination of the breeding ecology of garden birds (Chapter 3), the dawn chorus provides both a means to demonstrate ownership of a breeding territory and a mechanism to advertise your status as a mate. Research suggests that singing at dusk and, particularly, dawn may provide an honest signal of the energetic status of the singing male. If this is the case, then we might expect the availability of food in garden feeders to shape the performance of those males with access to it. Experimental work on Blackbirds supports this, with Cuthill & MacDonald (1990) finding that food-supplemented males sang significantly more than unsupplemented males. This difference was largely the result of supplemented males initiating song earlier and having higher peak rates of song delivery. Could this mean that male Blackbirds with access to a reliable food source at garden feeding stations are more likely to attract a mate than those nesting in other habitats where perhaps food is more limiting? A similar piece of work, this time on Great Tits, casts some uncertainty on whether the relationship is as simple as it appears from the work done on Blackbirds. Katja Saggese and her collaborators provided male Great Tits with a continuous food supply over two weeks and then compared their singing activity with a group of unfed males (Saggese et al., 2011). In contrast to Cuthill & MacDonald’s findings, the food supplemented males started their dawn singing later than the control males, an effect that still continued two weeks after the provision of food had ended. The researchers were unsure of the reasons for what they had observed, but they felt that it could have been due to the presence of predators, attracted to the feeding stations, or something about the quality of the food itself.

FIG 42. Male Blackbirds with access to supplementary food sing for longer than those without access to such food, suggesting that the food provided at garden feeding stations might help local birds in defence of their territories and mate attraction. (Mike Toms)
FOOD PROVISION AND MOVEMENTS
Birds tend to reduce the size of the area over which they forage when provisioned with supplementary food, something that can lead to a reduction in the size of breeding territory and even a change in mating system in the case of Dunnock (Davies & Lundberg, 1984). As we will see in a moment in relation to Blackcaps, the provision of food in gardens can have a significant impact on movement and wintering behaviour.
Although not garden-related, the scale of feeding impacts can be seen from work on the White Stork Ciconia ciconia, a species that was wholly migratory in Europe but which has established resident populations across parts of Iberia largely in response to the year-round food available at landfill sites (Gilbert et al., 2016). Landfills and food discarded by people have also played a role in the expansion of urban-breeding Lesser Black-backed Gulls Larus fuscus here in the UK (Coulson & Coulson, 2008), a species that now winters here in large numbers, when it formerly wintered in southern Europe and north and west Africa.
FOOD PROVISION AND THE EVOLUTION OF NEW BEHAVIOUR – WINTERING BLACKCAPS
The Blackcap is a common summer visitor to much of Britain, wintering in southern Europe and south into North Africa. Although there are occasional records, the Blackcap was rarely encountered in Britain during the winter months 60 years ago (Stafford, 1956). However, since the 1950s we have seen a rapid and substantial increase in wintering records, as information from both the Garden Bird Feeding Survey and Bird Atlas 2007–11 show (Balmer et al., 2013). Data from BTO-led bird atlases show that the Blackcap’s wintering range in the UK has expanded by 77 per cent over the last 30 years. The increase in wintering numbers has come about because Blackcaps breeding in southern Germany and Austria have increasingly migrated in a northwesterly direction to Britain for the winter, rather than in a southwesterly direction to traditional wintering areas located in southern Spain (Helbig et al., 1994). This new migration strategy has been shown to be genetically encoded (Berthold et al., 1992), and is maintained via reproductive isolation and assortative mating, linked to fitness benefits on the breeding grounds (Bearhop et al., 2005).
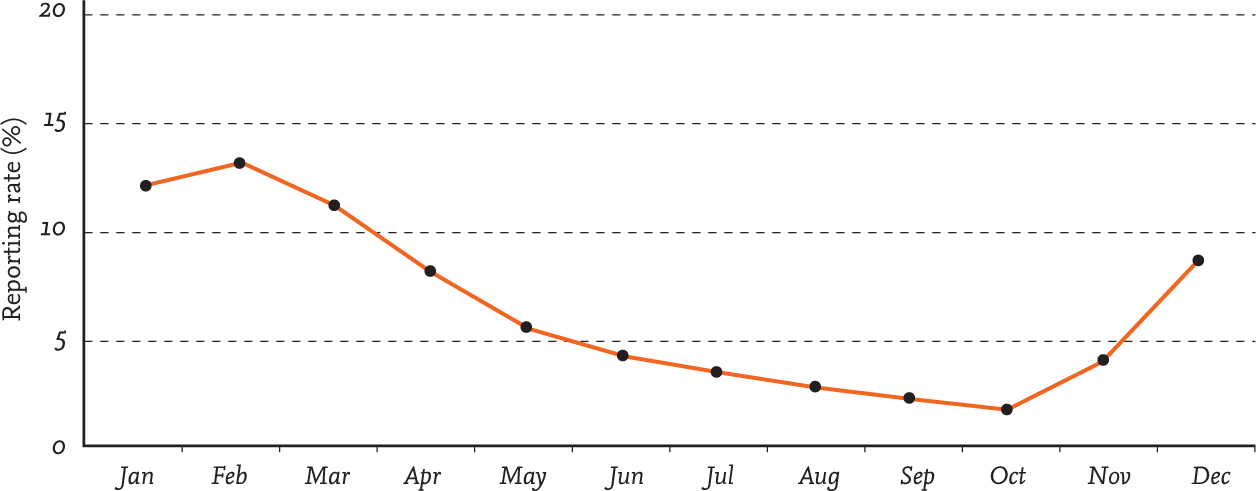
FIG 43. The use of gardens by Blackcaps peaks during the winter months. Data reproduced from BTO Garden BirdWatch with permission.
Early observations of increasing numbers of wintering Blackcaps coincided with the introduction of commercial bird foods (Callahan, 2014), suggesting that supplementary feeding at garden feeding stations might be applying a selection pressure for the evolution of this new migratory route. Interestingly, while the Blackcaps wintering in southern Spain are predominantly frugivorous in their dietary choices, those wintering in Britain are known to use a wide variety of supplementary food types (Tellería et al., 2013; Plummer et al., 2015). The availability of supplementary food is unlikely to be the sole driver of evolutionary change, however, and it is likely that a changing climate has also had some influence over the adoption of this new migration route and new wintering area. Winter conditions in Britain, and more widely across the Northern Hemisphere, are becoming milder, something that has enabled a number of species to shift their wintering range northwards (IPCC, 2013).
Unravelling these different drivers requires access to long-term datasets on food provision, Blackcap occurrence and climatic factors. Fortunately, BTO researchers have been able to use data from BTO’s weekly Garden BirdWatch survey to explore the question of whether food provision has played a part in the evolution of a new migratory strategy in this central European Blackcap population (Plummer et al., 2015). Earlier work using the BTO Garden BirdWatch dataset has revealed that Blackcaps are strongly associated with suburban gardens during the winter months (Chamberlain et al., 2004a) which makes this dataset ideal for examining the interactions between food provision and Blackcap wintering behaviour. Examination of the foods provided in Garden BirdWatch gardens and their use by wintering Blackcaps revealed a strong preference by the birds for sunflower hearts and fat-based products. By calculating an annual measure of the proportion of winter weeks in which these foodstuffs were provided at each of 3,806 sites – each of which had a minimum of at least 16 weekly submissions in a minimum of at least nine winters – it has been possible to explore the relationship between Blackcap occurrence and food provision and how this has changed over time.

