
Полная версия:
Эра млекопитающих: Из тени динозавров к мировому господству
Первой группой, с двумя отверстиями в черепе, были диапсиды. Они станут предками ящериц, змей, крокодилов, динозавров, птиц и черепах (у которых отверстия зарастут). Второй группой, с одним отверстием, были синапсиды. Они дадут ошеломляющее разнообразие видов, в том числе – через 100 с лишним миллионов лет – млекопитающих.
Это вымышленная история, и, вероятно, последовательность событий не была в точности такой. Правда в том, что около 325 млн лет назад – в тот период истории Земли, который называется пенсильванским (он же верхний каменноугольный), – существовала предковая популяция мелких чешуйчатых животных, обитавших в пышных заболоченных лесах, часто затапливаемых при подъеме уровня моря. Они разделились, и одна ветвь родословного древа привела к рептилиям, другая – к млекопитающим.
Откуда нам это известно? Палеонтологи – специалисты по древней жизни вроде меня – располагают двумя ключевыми видами свидетельств. Эти свидетельства я буду привлекать на протяжении всей книги, рассказывая историю эволюции млекопитающих.
Во-первых, у нас есть ископаемые остатки и окружающая их порода. Окаменелости – прямое подтверждение существования видов, которые когда-то были живыми; это подсказки, которые палеонтологи ищут по всему миру, нередко терпя зной, холод, сырость, дождь, безденежье, комаров, войны и другие трудности. Многие из нас чувствуют себя детективами, расследующими события отдаленных времен, и в этом смысле окаменелости аналогичны волоскам и отпечаткам пальцев, оставшимся на месте преступления. Они сообщают нам, кто, где и когда обитал, а порой ископаемые остатки могут поведать о доисторических драмах – о хищниках, терзающих добычу, о жертвах наводнений, об уцелевших после страшных вымираний. Наиболее известны ископаемые остатки тел – подлинные части когда-либо живших организмов: кости, зубы, раковины, листья. Бывают также следы жизнедеятельности ископаемых организмов – свидетельства их поведения или то, что они оставляют после себя: отпечатки лап, норки, яйца, следы зубов или копролиты (окаменелый помет).
Окаменелости не валяются на улице или в земле у нас на огороде – они заключены внутри пород, таких как песчаники и сланцы. Разные породы образуются в различных условиях, и некоторые можно датировать химическими методами: подсчитывается соотношение материнского и дочернего изотопов, а затем на основании известных скоростей радиоактивного распада элементов, полученных в лаборатории, вычисляется возраст породы. Все это обеспечивает важный контекст для понимания того, когда и в какой среде обитали ископаемые существа.
Второй тип свидетельств окружает нас повсюду, и, чтобы найти их, не нужно ни особого искусства, ни везения. Это ДНК, которую мы и все остальные организмы несем внутри своих клеток. ДНК – инструкция, которая делает нас теми, кто мы есть; генетический код, отвечающий за то, как мы выглядим, какова наша физиология, как мы развиваемся и как производим на свет будущие поколения. Вместе с тем ДНК – это архив; в миллиардах пар нуклеотидов, составляющих наш геном, записана эволюционная история. По мере того как со временем меняются виды, меняется и их ДНК. Гены мутируют, перемещаются, включаются и выключаются. Участки ДНК дублируются или выпадают. Вставляются новые фрагменты. Соответственно, когда общий предок разделяется на два вида, их ДНК постепенно накапливают все больше различий, поскольку каждый вид идет своим путем и приспосабливается к собственным меняющимся условиям. А значит, можно взять последовательности ДНК современных видов, сопоставить их и составить родословное древо, объединяя в группы виды с наиболее сходными ДНК. Есть еще один изящный фокус. Можно взять любые два вида, подсчитать количество различий в ДНК, а затем, имея представление о скорости изменений ДНК в лабораторных экспериментах, вычислить, когда эти виды отделились друг от друга.
Сочиняя свой рассказ о болоте, пострадавшем от наводнения, я использовал оба типа свидетельств. Исследования ДНК указывают, что линии рептилий и млекопитающих разделились около 325 млн лет назад. Ископаемые остатки и породы говорят нам о том, каким был этот мир – ландшафт, совсем непохожий на современный.
Карта Земли пенсильванского периода почти неузнаваема. На ней было всего два крупных материка – Гондвана с центром на Южном полюсе и охватывавшая экватор Лавразия, с востока обрамленная грядой островов. На протяжении многих миллионов лет Гондвана медленно, со скоростью, сравнимой, пожалуй, со скоростью роста наших ногтей, дрейфовала на север, пока не столкнулась с Лавразией. Так появилась Пангея, суперконтинент, на котором будут протекать ранние стадии эволюции динозавров и млекопитающих. При столкновении две литосферные плиты смялись и образовали протяженную гряду гор параллельно экватору. По масштабам она была сравнима с современными Гималаями. Нынешние скромные Аппалачи – остаток этой некогда величественной горной цепи.
Тропические и субтропические регионы по обе стороны экваториальной горной гряды изобиловали жизнью. Это были каменноугольные болота, названные так потому, что основная доля каменного угля, обеспечившего промышленную революцию – особенно добывавшегося в Европе, а также на востоке и на Среднем Западе США, – образовалась именно в этих болотах. Этот уголь состоит из отмерших, захороненных и спрессованных остатков гигантских быстрорастущих лепидодендронов и каламитов. Они ничуть не походили на пальмы, магнолии и дубы, которые ныне так часто встречаются в подобных буйных лесах. Более того, эти древние деревья не цвели, не давали плодов и орехов. Они были близкими родичами плаунов и хвощей, теперь изредка попадающихся в подлеске, – печальное напоминание об их былой славе. Пенсильванские деревья – вместе с гигантскими стрекозами, жужжавшими вокруг их ветвей, и многоножками, шуршавшими у подножия их стволов, – росли так быстро за счет того, что в воздухе тогда было намного больше кислорода, примерно на 70 % больше, чем в наши дни.
Деревья образовали обширные дождевые леса, которые прижимались к берегам мелководных морей, омывающих растущий суперконтинент, и множества впадавших в них рек, притоков, дельт и лиманов. Сверху эти болота, вероятно, выглядели как пойма Миссисипи в современной Луизиане: густой покров из сплетающихся друг с другом деревьев и растений поменьше – одни торчат на островках ила среди запутанной сети ручьев, другие тянутся ползучими корнями к воде, и повсюду карабкаются, прыгают и летают всевозможные создания. Но ни птиц, ни комаров, ни бобров, ни выдр, ни других покрытых мехом млекопитающих. Все они появятся намного позже, совсем в другом мире, хотя их предки действительно обитали в этих каменноугольных болотах.
Почему столько деревьев полегло и обратилось в уголь? Потому что болота постоянно затапливало. Уровень моря поднимался и падал в пульсирующем ритме. Пенсильванский мир был ледниковым, причем это было предпоследнее крупное оледенение (о последнем, когда царили мамонты и саблезубые тигры, мы расскажем позже). Замерзла не вся планета; разумеется, каменноугольные болота не замерзли. Но Южный полюс Гондваны и юг Пангеи покрывала огромная шапка ледников. Самим своим существованием она была обязана как раз каменноугольным болотам: растущие в таком количестве гигантские деревья поглощали из атмосферы углекислый газ, и с уменьшением количества этого парникового газа, изолирующего планету, температура резко упала. На протяжении десятков миллионов лет ледяная шапка то увеличивалась, то сокращалась, как бы управляя уровнем Мирового океана. Лед таял, моря наступали, заливая болота, деревья гибли и оказывались погребенными. Затем ледник снова рос, оттягивая воду из океана, уровень моря падал, и освобождалось место для распространения болот. Так и шло – туда-сюда. Мы знаем это благодаря тому, что пенсильванские отложения часто образуют полосчатые последовательности – так называемые циклотемы. Это повторяющиеся ряды тонких слоев, откладывавшихся то на суше, то в воде, с промежуточными включениями каменного угля.
Окаменелостей этого периода очень много, особенно в Северном Иллинойсе, где я вырос. Они залегают в циклотемах выше и ниже угольных слоев. Лучшие из них попадаются на берегах Мазон-Крика, тихого притока реки Иллинойс, и в карьерах к востоку. В пенсильванский период это было пограничье между морем и болотом, где обитателей дождевых лесов смывало в воду, они опускались на дно и оказывались замурованными в саркофагах из железняка – овальных, сплюснутых конкрециях цвета ржавчины, которые можно найти на дне ручья или в отвале карьера. Подростком я охотился за этими конкрециями у маленького городка Уилмингтона на 66-м шоссе, где росла моя мама. Я копался в отвалах давно заброшенных карьеров, которые больше века назад привлекли моих итальянских прадеда и прабабку обещанием новой жизни на Среднем Западе. Сложив конкреции в ведро, я относил их домой, выставлял во двор на суровые зимние холода Большого Чикаго, чтобы они то замерзали, то оттаивали при колебаниях температуры. Заметив, что конкреция начала трескаться, я довершал дело молотком.
Если мне везло, конкреция разламывалась, открывая сокровище: окаменелость на одной половине, ее отпечаток на другой. Всякий раз это был мистический опыт – знать, что ты первый, кто видит это создание – которое некогда было живым! – умершее более 300 млн лет назад. Во многих расколотых конкрециях оказывались растения: листья папоротника, кусочки коры каламитов, обломки корней лепидодендронов. Особенно мне нравились медузы, которых ветераны охоты за окаменелостями в Мазон-Крике пренебрежительно называли «лепешками», – и я всегда радовался, увидев креветку или червя.
Больше всего мне хотелось – но так и не повезло – отыскать тетрапода, наземное животное, обладающее скелетом. Из книг, которые я жадно глотал после школы и в тихие послеобеденные часы в выходные, я знал, что тетраподы произошли от рыб и выползли на сушу 390 млн лет назад, еще до пенсильванского периода. Эти первые четвероногие были амфибиями, и им все еще приходилось возвращаться в воду, чтобы отложить яйца. В Мазон-Крике даже находили кое-какие скелеты примитивных амфибий, отдаленных родичей лягушек и саламандр.
В какой-то момент пенсильванского периода от этих амфибий отделилась новая группа. Это были амниоты, более специализированные тетраподы, названные так по строению своих яйцеклеток, чьи внутренние мембраны (амниотические оболочки) окружали эмбрион, защищая его от высыхания. Новый тип яйцеклетки открывал огромные новые возможности: амниоты уже не были прикованы к воде и могли откладывать яйца на суше, а значит, осваивать новые рубежи. Вершины деревьев, норы, равнины, горы, пустыни. Лишь с появлением амниотического яйца тетраподы смогли по-настоящему покинуть море и покорить сушу.
Именно из амниот выделились линии рептилий и млекопитающих – диапсид и синапсид, – как двое детей одних и тех же родителей. Это не просто аналогия, именно так эволюция производит новые виды, семейства, классы. Виды непрерывно меняются под влиянием изменений окружающей среды – так работает дарвиновская эволюция путем естественного отбора. Иногда популяции одного вида оказываются в изоляции друг от друга, например из-за наводнения, пожара или подъема горной гряды. Каждая популяция продолжает меняться под воздействием естественного отбора, и если изоляция сохраняется достаточно долго, то каждая из них меняется по-своему, приспосабливаясь к различным обстоятельствам, настолько, что они уже не выглядят одинаково, не ведут себя одинаково и не могут спариваться друг с другом. Тогда вместо одного вида появляются два. Эти два вида могут снова разделиться – из двух получится четыре и т. д. Жизнь всегда диверсифицируется таким образом, разветвляясь подобно дереву возрастом в 4 млрд лет. Вот почему для наглядного представления родословных – как вымерших видов, так и человеческих семей – мы используем деревья, а не сети, дорожные карты, треугольники или иное графическое подспорье.
Разделение диапсид и синапсид – которое на самом деле началось с неприметного события, когда один маленький чешуйчатый предковый вид разделился на два, – стало одним из поворотных моментов в эволюции позвоночных. И я знал, что диапсиды и синапсиды – каждые со своей характерной, неповторимой системой отверстий в черепе и челюстных мышц – разошлись как раз примерно в то время, когда образовались конкреции Мазон-Крика. При каждом ударе молотка я надеялся найти Святой Грааль палеонтологии, который поможет рассказать эту историю, но, увы, так и не нашел.
Однако охотникам за окаменелостями из других регионов Северной Америки везло больше. Одно важное открытие было сделано в 1956 г., когда гарвардская полевая экспедиция под руководством легендарного палеонтолога Альфреда Ромера обследовала заброшенную угольную шахту во Флоренции (Новая Шотландия) у Атлантического побережья. Один из техников, Арни Льюис, заметил несколько окаменелых пней сигиллярии – дерева, родственного лепидодендрону, с раздвоенной кроной на верхушке, придававшей ему вид великанской кисточки. Пни сохраняли первоначальное положение, словно поднимающееся море затопило их только вчера, а не 310 млн лет назад – столько им было лет на самом деле. Пробираясь по узким штрекам затопленной шахты, экспедиции удалось набрести на пять таких пеньков. Заглянув в них, ученые обнаружили сюрприз: десятки окаменелых скелетов! Бедные животные, вероятно, пытались укрыться от наступающего моря в дуплах, не понимая, что найдут себе там могилу. Внутри одного дерева оказалось более двадцати особей, среди которых были амфибии, диапсиды и синапсиды: триада древнейших наземных тетрапод.
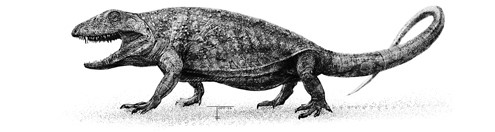
Archaeothyris.
Рисунок Тодда Маршалла
Синапсиды впоследствии были описаны как два новых вида, археотирис (Archaeothyris) и эхинерпетон (Echinerpeton). Описал их магистрант Роберт Райс, только что перебравшийся из Румынии в Канаду. Ныне один из ведущих палеонтологов мира, он набил себе руку как раз на этих древних синапсидах. Название Archaeothyris, «древнее окно», он выбрал, чтобы подчеркнуть самый важный признак этого животного: широкое, как иллюминатор, отверстие за глазницей, в котором крепились более крупные и мощные мышцы, смыкающие челюсти, чем у предшественников. Именно это одиночное отверстие, которое в анатомии называется боковым височным, определяет природу синапсид. У всех синапсид – от первопроходцев каменноугольных болот до современных слонов, землероек и летучих мышей – есть это отверстие или его видоизмененный вариант. Оно есть и у нас, и его можно ощутить при смыкании челюсти. Положите ладонь на скулу, кусните как следует, и вы почувствуете, как мышцы вашей щеки сокращаются. Эти мышцы проходят сквозь рудимент височного отверстия, которое у современных млекопитающих более или менее слилось с глазницей, но все еще участвует в креплении височных мышц, которые тянутся от виска к верхней части нижней челюсти, придавая силу укусу. Это одиночное отверстие появилось на раннем этапе истории синапсид, сразу после их отделения от диапсид, у которых развилось два таких заглазничных отверстия.
Если бы вы увидели, как археотирис пробирается через каменноугольное болото, он показался бы ничем не примечательным. Полметра в длину от морды до хвоста, с маленькой головкой на длинном узком туловище. О строении его конечностей известно недостаточно, но сохранившиеся кости не оставляют сомнений, что лапки у него были расставлены в стороны, как у ящерицы или крокодила. Он явно не был создан для быстрого бега. Однако при ближайшем рассмотрении становится ясно, что в некоторых отношениях он был исключительным. Мало того что в его черепе скрывались более мощные челюстные мышцы – в пасти у него был ряд изогнутых острых зубов. Один из передних зубов был заметно крупнее остальных и походил на миниатюрный клык. У амфибий, ящериц и крокодилов клыков нет[4]. Все эти животные обладают недифференцированными зубами, одинаковыми по всей длине челюсти. Однако у млекопитающих зубы гораздо разнообразнее: они делятся на резцы, клыки, предкоренные и коренные – такое разделение труда позволяет нам одновременно хватать, кусать и пережевывать. Полный набор зубов, свойственный млекопитающим, разовьется позже, в ходе многоступенчатой эволюции, но маленькие клыки археотириса – веяние зубной революции.

Два основных типа строения черепа наземных позвоночных: диапсиды с двумя заглазничными отверстиями для крепления челюстных мышц и синапсиды – включая человека – с одиночным отверстием. Отверстия отмечены стрелками.
Рисунок Сары Шелли
В комплекте эти крупные мышцы челюстей, острые зубы и клыки археотириса представляли собой арсенал оружия для охоты на крупных насекомых, а может быть, даже других тетрапод вроде эхинерпетона. Этот второй синапсид из Новой Шотландии с легкостью мог бы, свернувшись калачиком, уместиться на книжном развороте. Но его скудные ископаемые остатки демонстрируют одну необычную особенность, из-за которой он получил свое название «колючей рептилии». На шейных и спинных позвонках у него были шипы, вытянутые вверх длинными язычками. Выстроившись в ряд, они образовывали вдоль спины небольшой парус, который мог использоваться для бравады, или в качестве солнечной батареи, чтобы согреваться в холодные дни, или в качестве кондиционера, чтобы охлаждаться в жару, или еще для чего-нибудь.
Гораздо более знаменито другое ископаемое животное, у которого и парус на спине был гораздо больше: диметродон (Dimetrodon), живший в следующую эпоху после пенсильванской – в пермский период. Диметродона часто путают с динозаврами – его можно видеть рядом с тираннозавром на открытках, бок о бок с бронтозавром и стегозавром в наборах игрушек. Но он не динозавр, а синапсид. Точнее, разновидность примитивных синапсид – пеликозавр.
Пеликозавры были первой крупной эволюционной волной линии синапсид; они первыми диверсифицировались и распространились по растущему суперконтиненту Пангея. Кроме того, именно у них впервые стали появляться фирменные признаки, которые ныне, более 300 млн лет спустя, все еще отличают млекопитающих от амфибий, рептилий и птиц. Такие признаки, как височное отверстие для мышц и клыки, – признаки, которые мы уже наблюдали у археотириса и эхинерпетона. Эти два вида из Новой Шотландии и были древнейшими пеликозаврами, основателями первой великой династии на пути к диметродону, а в конечном итоге и к млекопитающим.
На закате пенсильванского периода пеликозавровые синапсиды обитали во всех экваториальных регионах Пангеи по обе стороны горного хребта, который еще продолжал расти. Одни питались насекомыми, другие – более мелкими тетраподами и рыбой, а некоторые начали экспериментировать с новой пищей, которой прежде пренебрегали, – листьями и стеблями. Они диверсифицировались, но оставались второстепенными элементами экосистем, где царили амфибии, без труда размножавшиеся во влажных каменноугольных лесах и потому процветавшие.
Затем, примерно 303–307 млн лет назад, мир резко изменился – наступил так называемый кризис каменноугольных лесов. Климат стал суше, планету бросало то в холод, то в жар, ледники таяли и в грядущем пермском периоде исчезли совсем. Каменноугольные болота приходили в упадок, так как каламитам, лепидодендронам и сигилляриям стало труднее расти в засушливых условиях. Их вытесняли хвойные, цикадовые и другие семенные растения, более засухоустойчивые. Постоянно влажные дождевые леса в тропиках уступили место полупустыням с более выраженной сменой времен года, а другие области Пангеи обратились в выжженные пустыни. Это отразилось в геологической летописи, в которой слои каменного угля и циклотемы внезапно сменяются «красными слоями» окисленного железа, образовавшегося в условиях сухого климата.
Эти изменения нанесли тяжелый удар по биоразнообразию. Больше всего пострадали растения. Мало того что пенсильванская флора каменноугольных болот сменилась более засухоустойчивыми семенными растениями, происходило массовое вымирание. Многие пенсильванские виды исчезли – одни не оставили ни потомков, ни близких родичей, от других остались лишь низкорослые неприметные собратья. Общим счетом вымерла половина всех семейств растений пенсильванского периода. В ископаемой летописи растений известно всего два массовых вымирания, и это одно из них. Второе случилось в конце пермского периода, и о нем мы вскоре еще расскажем. То есть кризис каменноугольных лесов с ботанической точки зрения был более катастрофичным, чем мел-палеогеновое столкновение с астероидом, погубившее динозавров.
Что стало с животными, обитавшими в этих лесах? На этот вопрос отвечает работа молодой исследовательницы Эммы Данн. Эмма, уроженка Ирландии, защитившая диссертацию в Англии, – олицетворение нового поколения палеонтологов. Как и легионы охотников за ископаемыми в прежние времена, она собирает окаменелости, но, кроме того, она специализируется на работе с массивами данных и современных статистических методах. Всегда есть соблазн раскрутить сенсационную историю на основании парочки новых находок, но представители поколения Эммы, желающие по-настоящему разобраться в закономерностях и процессах эволюции, мыслят как аналитики финансового рынка или инвестбанкиры. Они собирают тонны сведений, используют статистические модели с поправкой на неопределенность и проверяют гипотезы, сравнивая их убедительность, опираясь на цифры, а не на интуицию.
Исходя из этого, Эмма создала базу данных, включающую более тысячи находок тетрапод каменноугольного и пермского периодов, отмечая, к каким группам они принадлежат и где были найдены. Она применяла статистические методы, чтобы сгладить погрешности выборки, неизбежные в палеонтологических исследованиях, столь зависимых от случайных находок окаменелостей в немногих удачных местах, где кости и зубы сохранились через сотни миллионов лет. В итоге она построила статистические модели, чтобы проверить, как общее разнообразие и распределение видов – включая амфибий, диапсид и синапсид – менялись при исчезновении дождевых лесов.
Результаты обескураживали. На пограничье каменноугольного и пермского периодов наблюдался значительный спад разнообразия, многие из тетрапод каменноугольных лесов вымерли. По-видимому, все произошло не одномоментно, а длилось несколько миллионов лет, в ходе вытеснения тропических каменноугольных лесов полупустынями, надвигавшимися с запада на восток. Эта смена среды обитания – скорее, по-видимому, плавный переход, чем коллапс, – привела к появлению более открытых ландшафтов, благоприятных для миграций. Тетраподы, способные переносить сухой климат, теперь могли распространяться гораздо шире. Это были не амфибии, так долго господствовавшие в пенсильванском мире болот – ведь их репродуктивные стратегии приковывали их к воде. Но диапсиды и синапсиды обнаружили, что у них есть сверхспособность, идеально подходящая к новой реальности, – их амниотическое яйцо с мембранами, питающими эмбрион и оберегающими его от высыхания. Благодаря их свободному перемещению складывались связи между прежде изолированными областями, и таким образом формировались новые виды, новые типы строения тела, росли размеры, менялись рацион питания и модели поведения.
Когда на смену каменноугольным лесам пришли открытые полупустыни и пустыни, начался пермский период, и Земля стала планетой пеликозавров. Самый наглядный символ новой эры владычества пеликозавров – диметродон, тот знаменитый персонаж с парусом на спине, хорошо известный по десяткам скелетов, найденных в Техасе. Диметродона не зря так часто принимают за динозавра: его строение тела выглядит – как бы это выразиться? – рептильным. Массивный, неповоротливый, с длинным хвостом и острыми зубами, вряд ли он мог быстро бегать на своих коротких растопыренных лапах. Даже мозг у него был маленьким и трубкообразным, как у динозавров, а не как у млекопитающих, чей мозг крупнее и имеет сильно увеличенный складчатый мозжечок, что говорит о более развитом интеллекте и большей чувствительности. По этим признакам диметродон, вероятно, недалеко ушел от предковой популяции мелких чешуйчатых зверюшек, которая в пенсильванский период разделилась на диапсид и синапсид.

Пеликозавры – примитивные синапсидные предки млекопитающих: диметродон с парусом на спине (вверху) и пузатая растительноядная казеида.
Фотографии Х. Зелла и Райана Соммы соответственно
Однако в других отношениях диметродон заметно отличался от предков. Особенно это очевидно, если посмотреть на его пасть, где зубы совсем не похожи на однообразные ряды лезвий или столбиков большинства амфибий и диапсид. В передней части морды красовались большие закругленные резцы, за которыми следовали крупные клыки, а за ними, вдоль щеки, – ряд более мелких заклыковых зубов, острых и изогнутых. Это была следующая ступень в эволюции классической зубной системы млекопитающих, после развития клыков у ранних пеликозавров вроде найденных в пеньках археотирисов. Изменениям строения зубов соответствуют изменения челюстных мышц – они укрупняются и крепятся к более массивной и глубоко посаженной нижней челюсти, что еще больше увеличивает силу укуса. Изменения затронули и позвоночник – отдельные позвонки теперь соединяются так, чтобы ограничить неудобные волнообразные движения из стороны в сторону, столь характерные для рептилий и амфибий.

