 Полная версия
Полная версияEssays Upon Heredity and Kindred Biological Problems
But if the first segmentation spheres are not able to develope into a complete organism, how can this be the case with one of the later embryonic cells, or one of the cells of the fully developed animal body? It is true that we speak of certain cells as being ‘of embryonic character,’ and only recently Kölliker125 has given a list of such cells, among which he includes osteoblasts, cartilage cells, lymph corpuscles, and connective tissue corpuscles: but even if these cells really deserve such a designation, no explanation of the formation of germ-cells is afforded, for the idioplasm of the latter must be widely different from that of the former.
It is an error to suppose that we gain any further insight into the formation of germ-cells by referring to these cells of so-called ‘embryonic character,’ which are contained in the body of the mature organism. It is of course well known that many cells are characterized by very sharply defined histological differentiation, while others are but slightly differentiated; but it is as difficult to imagine that germ-cells can arise from the latter as from the former. Both classes of cells contain idioplasm with a structure different from that which is contained in the germ-cell, and we have no right to assume that any of them can form germ-cells until it is proved that somatic idioplasm is capable of undergoing re-transformation into germ-idioplasm.
The same argument applies to the cells of the embryo itself, and it therefore follows that those instances of early separation of sexual from somatic cells, upon which I have often insisted as indicating the continuity of the germ-plasm, do not now appear to be of such conclusive importance as at the time when we were not sure about the localization of the idioplasm in the nuclei. In the great majority of cases the germ-cells are not separated at the beginning of embryonic development, but only in some one of the later stages. A single exception is found in the pole-cells (‘Polzellen’) of Diptera, as was shown many years ago by Robin126 and myself127. These are the first cells formed in the egg, and according to the later observations of Metschnikoff128 and Balbiani129, they become the sexual glands of the embryo. Here therefore the germ-plasm maintains a true unbroken continuity. The nucleus of the egg-cell directly gives rise to the nuclei of the pole-cells, and there is every reason to believe that the latter receive unchanged a portion of the idioplasm of the former, and with it the tendencies of heredity. But in all other cases the germ-cells arise by division from some of the later embryonic cells, and as these belong to a more advanced ontogenetic stage in the development of the idioplasm, we can only conclude that continuity is maintained, by assuming (as I do) that a small part of the germ-plasm persists unchanged during the division of the segmentation nucleus and remains mixed with the idioplasm of a certain series of cells, and that the formation of true germ-cells is brought about at a certain point in the series by the appearance of cells in which the germ-plasm becomes predominant. But if we accept this hypothesis it does not make any difference, theoretically, whether the germ-plasm becomes predominant in the third, tenth, hundredth, or millionth generation of cells. It therefore follows that cases of early separation of the germ-cells afford no proof of a direct persistence of the parent germ-cells in those of the offspring; for a cell the offspring of which become partly somatic and partly germ-cells cannot itself have the characters of a germ-cell; but it may nevertheless contain germ-idioplasm, and may thus transfer the substance which forms the basis of heredity from the germ of the parent to that of the offspring.
If we are unwilling to accept this hypothesis, nothing remains but to credit the idioplasm of each successive ontogenetic stage with a capability of re-transformation into the first stage. Strasburger accepts this view; and he believes that the idioplasm of the nuclei changes during the course of ontogeny, but returns to the condition of the first stage of the germ, at its close. But the rule of probability is against such a suggestion. Suppose, for instance, that the idioplasm of the germ-cell is characterized by ten different qualities, each of which may be arranged relatively to the others in two different ways, then the probability in favour of any given combination would be represented by the fraction (1/2)10 = 1/1024: that is to say, the re-transformation of somatic idioplasm into germ-plasm will occur once in 1024 times, and it is therefore impossible for such re-transformation to become the rule. It is also obvious that the complex structure of the germ-plasm which potentially contains, with the likeness of a faithful portrait, the whole individuality of the parent, cannot be represented by only ten characters, but that there must be an immensely greater number; it is also obvious that the possibilities of the arrangement of single characters must be assumed to be much larger than two; so that we get the formula (1/p), where p represents the possibilities, and n the characters. Thus if n and p are but slightly larger than we assumed above, the probabilities become so slight as to altogether exclude the hypothesis of a re-transformation of somatic idioplasm into germ-plasm.
It may be objected that such re-transformation is much more probable in the case of those germ-cells which separate early from the somatic cells. Nothing can in fact be urged against the possibility that the idioplasm of (e. g.) the third generation of cells may pass back into the condition of the idioplasm of the germ-cell; although of course the mere possibility does not prove the fact. But there are not many cases in which the sexual cells are separated so early as the third generation: and it is very rare for them to separate at any time during the true segmentation of the egg. In Daphnidae (Moina) separation occurs in the fifth stage of segmentation130, and although this is unusually early it does not happen until the idioplasm has changed its molecular structure six times. In Sagitta131 the separation does not take place until the archenteron is being formed, and this is after several hundred embryonic cells have been produced, and thus after the germ-plasm has changed its molecular structure ten or more times. But in most cases, separation takes place at a much later stage; thus in Hydroids it does not happen until after hundreds or thousands of cell-generations have been passed through; and the same fact holds in the higher plants, where the production of germ-cells frequently occurs at the end of ontogeny. In such cases the probability of a re-transformation of somatic idioplasm into germ-plasm becomes infinitely small.
It is true that these considerations only refer to a rapid and sudden re-transformation of the idioplasm. If it could be proved that development is not merely in appearance but in reality a cyclical process, then nothing could be urged against the occurrence of re-transformation. It has been recently maintained by Minot132 that all development is cyclical, but this is obviously incorrect, for Nägeli has already shown that direct non-cyclical courses of development exist, or at all events courses in which the earliest condition is not repeated at the close of development. The phyletic development of the whole organic world clearly illustrates a development of the latter kind; for although we may assume that organic development is not nearly concluded, it is nevertheless safe to predict that it will never revert to its original starting-point, by backward development over the same course as that which it has already traversed. No one can believe that existing Phanerogams will ever, in the future history of the world, retrace all the stages of phyletic development in precise inverse order, and thus return to the form of unicellular Algae or Monera; or that existing placental mammals will develope into Marsupialia, Monotremata, mammal-like reptiles, and the lower vertebrate forms, into worms and finally into Monera. But how can a course of development, which seems to be impossible in phylogeny, occur as the regular method of ontogeny? And quite apart from the question of possibility, we have to ask for proofs of the actual occurrence of cyclical development. Such a proof would be afforded if it could be shown that the nucleoplasm of those somatic cells which (e.g. in Hydroids) are transformed into germ-cells passes backwards through many stages of development into the nucleoplasm of the germ-cell. It is true that we can only recognise differences in the structure of the idioplasm by its effects upon the cell-body, but no effects are produced which indicate that such backward development takes place. Since the course of onward development is compelled to pass through the numerous stages which are implied in segmentation and the subsequent building-up of the embryo, etc., it is quite impossible to assume that backward development would take place suddenly. It would be at least necessary to suppose that the cells of embryonic character, which are said to be transformed into primitive germ-cells, must pass back through at any rate the main phases of their ontogeny. A sudden transformation of the nucleoplasm of a somatic cell into that of a germ-cell would be almost as incredible as the transformation of a mammal into an amoeba; and yet we are compelled to admit that the transformation must be sudden, for no trace of such retrogressive stages of development can be seen. If the appearance of the whole cell gives us any knowledge as to the structure of its nuclear idioplasm, we may be sure that the development of a primitive germ-cell proceeds without a break, from the moment of its first recognizable formation, to the ultimate production of distinct male or female sexual cells.
I am well aware that Strasburger has stated that, in the ultimate maturation of the sexual cells, the substance of the nuclei returns to a condition similar to that which existed at the beginning of ontogenetic development; still such a statement is no proof, but only an assumption made to support a theory. I am also aware that Nussbaum and others believe that, in the formation of spermatozoa in higher animals, a backward development sets in at a certain stage; but even if this interpretation be correct, such backward development would only lead as far as the primitive germ-cell, and would afford no explanation of the further transformation of the idioplasm of this cell into germ-plasm. But this latter transformation is just the point which most needs proof upon any theory except the one which assumes that the primitive germ-cell still contains unchanged germ-plasm. Every attempt to render probable such a re-transformation of somatic nucleoplasm into germ-plasm breaks down before the facts known of the Hydroids, in which only certain cells in the body, out of the numerous so-called embryonic cells, are capable of becoming primitive germ-cells, while the rest do not possess this power.
I must therefore consider as erroneous the hypothesis which assumes that the somatic nucleoplasm may be transformed into germ-plasm. Such a view may be called ‘the hypothesis of the cyclical development of the germ-plasm.’
Nägeli has tried to support such an hypothesis on phyletic grounds. He believes that phyletic development follows from an extremely slow but steady change in the idioplasm, in the direction of greater complexity, and that such changes only become visible periodically. He believes that the passage from one phyletic stage to another is chiefly due to the fact that ‘in any ontogeny, the very last structural change upon which the separation of germs depends, takes place in a higher stage, one or more cell-generations later’ than it occurred in a lower stage. ‘The last structural change itself remains the same, while the series of structural changes immediately preceding it is increased.’ I believe that Nägeli, being a botanist, has been too greatly influenced by the phenomena of plant-life. It is certainly true that in plants, and especially in the higher forms, the germ-cells only make their appearance, as it were, at the end of ontogeny; but facts such as these do not hold in the animal kingdom: at any rate they are not true in the great majority of cases. In animals, as I have already mentioned several times, the germ-cells are separated from the somatic cells during embryonic development, sometimes even at its very commencement; and it is obvious that this latter is the original, phyletically oldest, mode of formation. The facts at our disposal indicate that the germ-cells only appear, for the first time, after embryological development, in those cases where the formation of asexually produced colonies takes place, either with or without alternation of generations; or in cases where alternation of generations occurs without the formation of such colonies. In a colony of polypes, the germ-cells are produced by the later generations, and not by the founder of the colony which was developed from an egg. This is also true of the colonies of Siphonophora, and the germ-cells appear to arise very late in certain instances of protracted metamorphosis (Echinodermata), but on the other hand, they arise during the embryonic development of other forms (Insecta) which also undergo metamorphosis. It is obvious that the phyletic development of colonies or stocks must have succeeded that of single individuals, and that the formation of germ-cells in the latter must therefore represent the original method. Thus the germ-cells originally arose at the beginning of ontogeny and not at its close, when the somatic cells are formed.
This statement is especially supported by the history of certain lower plants, or at any rate chlorophyll-containing organisms, and I think that these forms supply an admirable illustration of my theory as to the phyletic origin of germ-cells, as explained in my earlier papers upon the same subject.
The phyletic origin of germ-cells obviously coincides with the differentiation of the first multicellular organisms by division of labour133. If we desire to investigate the relation between germ-cells and somatic cells, we must not only consider the highly developed and strongly differentiated multicellular organisms, but we must also turn our attention to those simpler forms in which phyletic transitions are represented. In addition to solitary unicellular organisms, we know of others living in colonies of which the constituent units or cells (each of them equivalent to a unicellular organism) are morphologically and physiologically identical. Each unit feeds, moves, and under certain circumstances is capable of reproducing itself, and of thus forming a new colony by repeated division. The genus Pandorina (Fig. I), belonging to the natural order Volvocineae, represents such ‘homoplastid’ (Götte) organisms. It forms a spherical colony composed of ciliated cells, all of which are exactly alike: they are embedded in a colourless gelatinous mass. Each cell contains chlorophyll, and possesses a red eye-spot, and a pulsating vacuole. These colonies are propagated by the sexual and asexual (Fig. II) methods alternately, although in the former case the conjugating swarm-cells cannot be distinguished with certainty as male or female. In both kinds of reproduction, each cell in the colony acts as a reproductive cell; in fact, it behaves exactly like a unicellular organism.
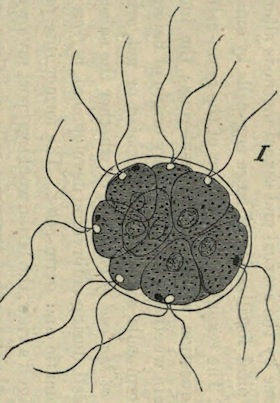
I. Pandorina morum (after Pringsheim), a swarming colony.
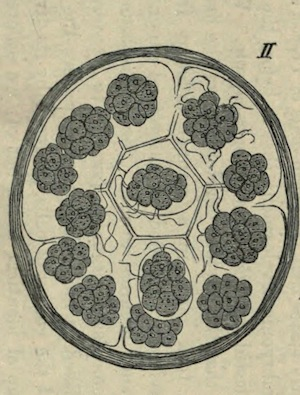
II. A colony divided into sixteen daughter colonies: all the cells alike.
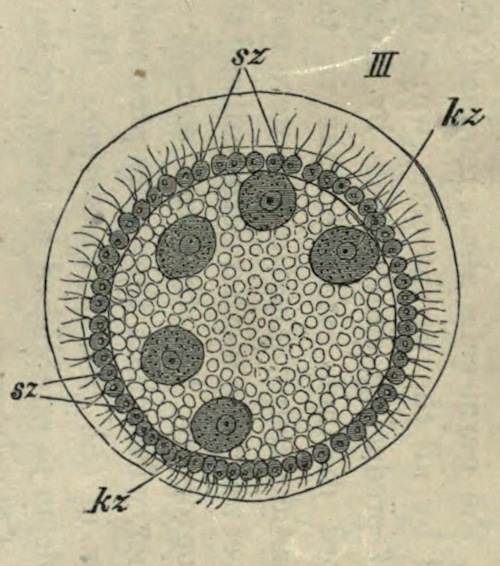
III. A young individual of Volvox minor (after Stein), still enclosed in the wall of the cell from which it has been parthenogenetically produced. The constituent cells are divided into somatic (sz), germ-cells (kz).
It is very interesting to find in another genus belonging to the same natural order, that the transition from the homoplastid to the heteroplastid condition, and the separation into somatic and reproductive cells, have taken place. In Volvox (Fig. III) the spherical colony consists of two kinds of cells, viz. of very numerous small ciliated cells, and of a much smaller number of large germ-cells without cilia. The latter alone possess the power of producing a new colony, and this takes place by the asexual and sexual methods alternately: in the latter a typical fertilization of large egg-cells by small spermatozoa occurs. The sexual differentiation of the germ-cells is not material to the question we are now considering; the important point is to ascertain whether here, at the very origin of heteroplastid organisms, the germ-cells, sexually differentiated or not, arise from the somatic cells at the end of ontogeny, or whether the substance of the parent germ-cell, during embryonic development, is from the first separated into somatic and germ-cells. The former interpretation would support Nägeli’s view, the latter would support my own. But Kirchner134 distinctly states that the germ-cells of Volvox are differentiated during embryonic development, that is, before the escape of the young heteroplastid organism from the egg-capsule. We cannot therefore imagine that the phyletic development of the first heteroplastid organism took place in a manner different from that which I have previously advocated on theoretical grounds, before this striking instance occurred to me. The germ-plasm (nucleoplasm) of some homoplastid organism (similar to Pandorina) must have become modified in molecular structure during the course of phylogeny, so that the colony of cells produced by its division was no longer made up of identical units, but of two different kinds. After this separation, the germ-cells alone retained the power of reproduction possessed by all the parent cells, while the rest only retained the power of producing similar cells by division. Thus Volvox seems to afford distinct evidence that in the phyletic origin of the heteroplastid groups, somatic cells were not, as Nägeli supposes, intercalated between the mother germ-cell and the daughter germ-cells in each ontogeny, but that the somatic cells arose directly from the former, with which they were previously identical, as they are even now in the case of Pandorina. Thus the continuity of the germ-plasm is established at least for the beginning of the phyletic series of development.
The fact, already often mentioned, that in most higher organisms the separation of germ-cells takes place later, and often very late, at the end of the whole ontogeny, proves that the time at which this separation of the two kinds of cells took place, must have been gradually changed. In this respect the well-established instances of early separation are of great value, because they serve to connect the extreme cases. It is quite impossible to maintain that the germ-cells of Hydroids or of the higher plants, exist from the time of embryonic development, as indifferent cells, which cannot be distinguished from others, and which are only differentiated at a later period. Such a view is contradicted by the simplest mathematical consideration; for it is obvious that none of the relatively few cells of the embryo can be excluded from the enormous increase by division, which must take place in order to produce the large number of daughter individuals which form a colony of polypes. It is therefore clear that all the cells of the embryo must for a long time act as somatic cells, and none of them can be reserved as germ-cells and nothing else: this conclusion is moreover confirmed by direct observation. The sexual bud of a Coryne arises at a part of the Polype which does not in any way differ from surrounding areas, the body wall being uniformly made up of two single layers of cells, the one forming the ectoderm and the other the endoderm. Rapid growth then takes place at a single spot, and some of the young cells thus produced are transformed into germ-cells, which did not previously exist as separate cells.
Strictly speaking I have therefore fallen into an inaccuracy in maintaining (in former works) that the germ-cells are themselves immortal; they only contain the undying part of the organism—the germ-plasm; and although this substance is, as far as we know, invariably surrounded by a cell-body, it does not always control the latter, and thus confer upon it the character of a germ-cell. But this admission does not materially change our view of the whole subject. We may still contrast the germ-cells, as the undying part of the Metazoan body, with the perishable somatic cells. If the nature and the character of a cell is determined by the substance of the nucleus and not by the cell-body, then the immortality of the germ-cells is preserved, although only the nuclear substance passes uninterruptedly from one generation to another.
G. Jäger135 was the first to state that the body in the higher organisms is made up of two kinds of cells, viz., ontogenetic and phyletic cells, and that the latter, the reproductive cells, are not a product of the former (the body-cells), but that they arise directly from the parent germ-cell. He assumed that the formation of germ-cells takes place at the earliest stage of embryonic life, and he thus believed the connexion between the germ-plasm of the parent and of the offspring had received a satisfactory explanation. As I have previously mentioned in the introduction, Nussbaum also brought forward this hypothesis at a later period, and also based it upon a continuity of the germ-cells. He assumed that the fertilized egg is divided into the cells of the individual and into the cells which effect the preservation of the species, and he supported this view by referring to the few known cases of early separation of the sexual cells. He even maintained this hypothesis when I had proved in my investigations on Hydromedusae that the sexual cells are not always separated from the somatic cells during embryonic development, but often at a far later period. Not only is the hypothesis of a direct connexion between the germ-cells of the offspring and parent broken down by the facts known in the Hydroids, and in the Phanerogams136 which resemble them in this respect, but even the instances of early separated germ-cells quoted by Jäger and Nussbaum do not as a matter of fact support their hypothesis. Among existing organisms it is extremely rare for the germ-cells to arise directly from the parent egg-cell (as in Diptera). If, however, the germ-cells are separated only a few cell-generations later, the postulated continuity breaks down; for an embryonic cell, of which the offspring are partly germ-cells and partly somatic cells, cannot itself possess the nature of a germ-cell, and its idioplasm cannot be identical with that of the parent germ-cell. In order to prove this, it is only necessary to refer to the arguments as to the ontogenetic stages of the idioplasm. In the above-mentioned instances, the continuity from the germ-substance of the parent to that of the offspring can only be explained by the supposition that the somatic nucleoplasm still contains some unchanged germ-plasm. I believe that the fundamental idea of Jäger and Nussbaum is quite correct: it is the same idea which has led me to the hypothesis of the continuity of the germ-plasm, viz., the conviction that heredity can only be understood by means of such an hypothesis. But both these writers have worked out the idea in the form of an hypothesis which does not correspond with the facts. That this is the case is also shown by the following words of Nussbaum—‘the cell-material of the individual (somatic cells) can never produce a single sexual cell.’ Such production undoubtedly takes place, not only in Hydroids and Phanerogams, but in many other instances. The germ-cells cannot indeed be produced by any indifferent cell of embryonic character, but by certain cells, and under circumstances which allow us to positively conclude that they have been predestined for this purpose from the beginning. In other words, the cells in question contain germ-plasm, and this alone enables them to become germ-cells.

