
Полная версия:
Боль в пояснице
Представители ортодоксальной медицины всегда оперируют официальными определениями заболеваний. Да, на практике в плане коммуникации врачи часто вынужденно «опускаются» до уровня обывателя и оперируют терминами, семантически доступными уму среднестатистического пациента: ВСД, остеохондроз и т.п. Понятно, что речь идет о словесной «двойной бухгалтерии»: два пишем три в уме. Пациенту озвучили одно, в карточку записали «полновесный» диагноз. Альтернативно одаренные «специалисты» или сознательно искажают официальные термины или вообще используют свои собственные. Выдуманные.
Ортодоксальные врачи – это материалисты. Озвученное врачом можно увидеть (МРТ, КТ, УЗИ, ЭНМГ, Доплер и т.п.), точно измерить (лабораторные методы диагностики), выстроить теорию (ретроспективные исследования как источник данных), спрогнозировать результат или подтвердить гипотезу (проспективные исследования). Скукота! То ли дело альтернативные… Они окутаны ореолом таинственности и, как правило, обещают «излечить» заболевания, при которых официальная медицина бессильна.
В официальной медицине патогенез заболевания рассматривается на различных уровнях – от молекулярных изменений, до нарушений в работе организма в целом. При этом выделяют специфические (зависящие от этиологии) и неспецифические (генетически обусловленные) механизмы заболевания. В зависимости от периодов подбирается тип лечения:
Этиотропная терапия направлена на устранение причины заболевания;
Патогенетическая терапия направлена на механизмы развития заболевания;
Симптоматическая (паллиативная) терапия применяется для устранения отдельных симптомов заболевания в дополнение к этиотропной и патогенетической.
Что же касается представителей альтернативной медицины, то об этиологии и патогенезе заболеваний они имеют смутное представление, если имеют вообще какое-либо. Закупорка энергетических каналов, потускнение чакр, меридианы там всякие, фасциальные поезда – это, конечно, все заумно, но это так не работает. Все, что касается доказательной медицины, у альтернативщиков вызывает неадекватные реакции, иногда переходящие в приступы неконтролируемой агрессии. Врачи, промышляющие халтурой или зарабатывающие медицинским лохотроном, всегда будут косо смотреть на доказательную медицину, как и специалисты, оказывающие реабилитационные услуги в области комплементарной медицины.
Пациенты также недолюбливают врачей, которые в своей работе предпочитают опираться на доказательную медицину. Такие врачи, по мнению пациентов, лишают их надежды на чудо исцеления. Правда, речь всегда почему-то идет о внешнем источнике исцеления и пассивной роли самого пациента.
Подтверждая взгляды [и опасения] Светличной, я слегка займусь «медикализацией» и постараюсь сделать так, чтобы в очередной раз «в массовое сознание произошло проникновение медицинского языка и стиля мышления, медицинских концепций и представлений о причинах, формах протекания и лечения болезней», другими словами [Ханс Баер и др.], по мере моих сил и знаний «навяжу людям биомедицинские нормы, ценности и представления», так сказать, слегка «погегемоню» по-биомедицински. И «прикроюсь» я особым статусом научной медицины, основанной на доказательствах.
Часто от пациентов можно слышать жалобы на врачей, что они толком не могут объяснить причину заболевания и как с ней справиться, и почему состояние только ухудшается. Некоторые специалисты вообще ничего не объясняют [потому что руководствуются здравым принципом: «дураки – это интеллектуальное большинство общества» и нет смысла вообще что–либо объяснять, поскольку сказанное будет или неправильно проинтерпретировано или сознательно перекручено]. Или же что разные врачи толкуют все по-своему, часто опровергая сказанное предыдущим врачом и, что их, пациентов, это еще больше запутывает и сбивает с толку… Претензия вполне понятна. Но читатель должен четко уяснить, что многие вещи простыми словами объяснить невозможно в принципе в силу их сложности. Мы не живем в эпоху Средневековья, и многие процессы достаточно хорошо изучены и описаны. Но они описаны научным языком и находу переводить с научного на «язык простолюдинов» – задача сложная, неблагодарная и не под силу большинству специалистов. [А метать бисер перед свиньями – то еще удовольствие].
Кардинальные отличия в представлениях о патогенезе заболеваний проще всего продемонстрировать на примере неспецифических болей в спине. Здесь воочию легко увидеть и понять принципиальную разницу между альтернативной и ортодоксальной медициной [и реабилитацией].
Желающие могут самостоятельно попробовать на пальцах объяснить своим близким/родным/коллегами/да кому угодно то, что я изложу ниже. Я слегка пройдусь по верхам, не вдаваясь в детали и не опускаясь на глубину.
Данная книга – это, в некоторой степени, попытка удовлетворить потребность любознательных пациентов знать причины болей в спине с академической точки зрения через призму реабилитации [поправку на ваши интеллектуальные способности делать я не буду].
Но все по порядку. Сначала – знакомство с основами, потом – в разрезе причин дегенеративных поражений позвоночника, их патогенеза, лечения, профилактики и реабилитации.
Механотрансдукция
Начну с фундамента официальной физической реабилитации на основе медицинских (ортодоксальных) представлений о боли в спине.
Международное название специалистов по физической реабилитации – «Физический терапевт». Не путать с врачебной специальностью «Врач физиотерапевт». С целью избежать путаницы я буду использовать термины «физическая реабилитация» и «специалист по физической реабилитации». «Реабилитологи» после 3-х месячных платных курсов не имеют никакого отношения к специалистам по физической реабилитации. Просто схожее название, вводящее людей в заблуждение.
Физическая реабилитация является частью механотерапии как производной от механобиологии. В основе механотерапии лежит использование механотрансдукции.
Механотрансдукция – это процесс преобразования механических стимулов в клеточный и молекулярный ответ. Другими словами – это механизм, с помощью которого механочувствительные клетки организма реагируют на нагрузку и преобразуют механические стимулы в биохимические клеточные реакции. Поэтому с помощью реабилитационных вмешательств (механических сигналов) через активацию определенных клеточных сигнальных путей возможно напрямую влиять на активацию генов, соответственно на пролиферацию (деление), дифференцировку, жизненный цикл и функционирование клеток.
Механотерапия – это «любое вмешательство, которое использует механические силы с целью изменения молекулярных путей и индукции клеточного ответа, усиливающего рост, моделирование, ремоделирование или восстановление тканей» [определение W.R. Thompson и соавт., 2016].
Почти все клетки организма так или иначе отвечают на механический стимул/стресс. Было идентифицировано множество типов клеток, способных воспринимать механические раздражители и реагировать на них: остеоциты, хондроциты, миобласты, фибробласты, кератиноциты и, что критически важно, – мезенхимальные стволовые клетки. Для реабилитации это имеет первостепенное значение, поскольку после травмы именно адекватная физическая нагрузка будет определять, как будет протекать пролиферация (деление/размножение) клеток и их дифференцировка (или/или/или). В зависимости от силы и длительности стимула будет зависеть, пойдет ли дифференцировка клеток по пути нейрогенеза, адипогенеза, хондрогенеза, миогенеза, фиброгенеза или остеогенеза. При неправильном лечении и реабилитации вместо мышечной ткани часто образуется жировая и фиброзная, вместо костной – хрящевая. Соответственно, с дальнейшим снижением функциональных возможностей поврежденного органа.
К механическим стимулам относятся: растяжение, сжатие, сдвиг, торсия (скручивание), гидростатическое давление, вибрация и движение жидкости параллельно клеточной мембране.
Для «считывания» сигналов в липидных слоях клеточных мембран располагаются различные типы механочувствительных рецепторов: ионные каналы, активируемые растяжением; интегрины; рецепторы факторов роста и рецепторы, связанные с G-белком.
Ионные каналы, активируемые растяжением, представляют собой трансмембранные белки, которые создают в клеточной мембране поры, достаточно большие для пропускания кальция и других катионов при их открытии. Механически вызванное натяжение/деформация мембраны способны открывать каналы, обеспечивая приток и отток ионов через клеточную мембрану в зависимости от внутреннего/внешнего градиента концентрации. Ионный баланс важен для многих клеточных функций и способствует жестко регулируемому электрическому потенциалу клеточной мембраны. Внутриклеточные концентрации кальция регулируют внутриклеточную передачу сигналов, полимеризацию актина (внутриклеточный механический сигналинг), ремоделирование цитоскелета и подвижность клеток.
Рецепторы, связанные с G-белком (GPCR), представляют собой крупные белки с 7 трансмембранными доменами. Внеклеточная часть связывает многие эффекторные белки и молекулы, такие как факторы роста, воспалительные цитокины, нейропептиды и гормоны, создавая конформационные изменения в структуре белка. После активации цитозольная (внутренняя) часть рецептора взаимодействует с внутриклеточными G-белками, воздействуя на сигнальный каскад в соответствии со специфичностью сигнала. Эти рецепторы также могут активироваться механическими нагрузками на поверхности клетки, которые инициируют каскады вторичных мессенджеров внутри клетки.
Рецепторы факторов роста активируются путем связывания с внеклеточными факторами роста, которые, в свою очередь, дополнительно активируют несколько опосредованных путей вторичных мессенджеров внутри клетки. Рецепторы факторов роста перекрестно связаны с другими механочувствительными рецепторами клеточной мембраны и могут усиливать их сигнальные эффекты.
Интегрины управляют клеточным сигналингом посредством актино-миозиновых связей между экстраклеточным матриксом, базальной (плазматической) мембраной и мембранными рецепторами ядра клетки.
Мембранные механочувствительные рецепторы могут воспринимать как механические, так и биохимические сигналы (гормоны, цитокины, ростовые факторы и пр.).
Механический сигнал передается по оси «матрикс – рецепторы/интегрины – цитоскелет – ядро».
Активированный рецептор на поверхности клеточной мембраны посредством последующей активации цитозольных медиаторов «каскадно» вовлекает в биохимическую реакцию транскрипционные факторы (исполнители команд), которые смещаются в ядро клетки, «включают» экспрессию генов, и, таким образом, запускают процесс синтеза необходимых аминокислот с последующей их сборкой в активные белки.
Итак. Трансформация внешнего сигнала (химического, электрического или механического) в клеточный ответ производится за счет «сигналинга» – последовательных биохимических реакций внутри клетки. Большинство транскрипционных ответов на нагрузки опосредуются через каскады вторичных мессенджеров и могут создавать длительные биологические эффекты. Клетка «запоминает» команды и «обучается» реакции на стимул.
Поскольку существует множество различных механочувствительных рецепторов, также существует множество сигнальных путей, которые клетка использует для генерации биохимического ответа.
Перечислю основные «сигнальные пути» и их краткую характеристику. Это понадобиться для понимания причин заболевания позвоночника, роли профилактики и реабилитации, а также влияет на выбор препаратов при лечении и для профилактики:
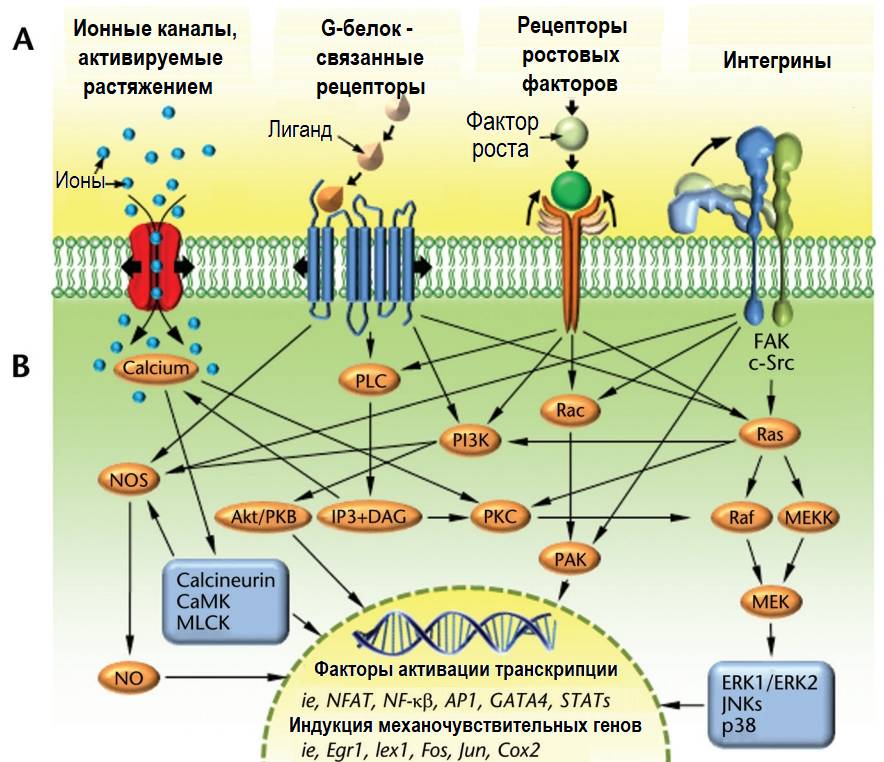
Механочувствительные каналы, рецепторы, сигнальные пути и факторы транскрипции. [исключительно в ознакомительных целях] (Thompson W. R.)
Сигнальные пути
Для удобства клеточный «сигналинг» буду описывать с поправкой на клетки скелетно-мышечной (опорно-двигательной) системы. Свое название сигнальные пути получили по названию ведущего белка или процесса.
Сигнальный путь Аденозинмонофосфат (АМФ)-активированной протеинкиназы (АМРК). АМФК – фермент, действующий как «энергетический сенсор», регулирующий энергетический гомеостаз при понижении уровня клеточного аденозинтрифосфата (АТФ). Участвует в катаболических и анаболических процессах на молекулярном и клеточном уровнях. Влияет на регуляцию аппетита в гипоталамусе.
[Далее: протеинкиназы – это большая группа ферментов, которые катализирует перенос концевого остатка фосфата с АТФ на различные группы в структуре белка; по сути – активаторы биохимических реакций].
В ходе реакции, катализируемой протеинкиназами, нейтральная спиртовая группа белка превращается в сложный эфир, несущий большой отрицательный заряд. Это приводит к значительным изменениям в структуре белка, сопровождается его структурными перестройками и изменением свойств. Химическая модификация молекулы белка, происходящая под действием протеинкиназ, является эффективным способом регуляции активности многих ферментов и других внутриклеточных белков. Остаток фосфорной кислоты, перенесенный протеинкиназой на спиртовую группу белка, может быть удален под действием другого фермента – фосфатазы. Таким образом, протеинкиназы и фосфатазы образуют две группы ферментов-антагонистов, способных осуществлять обратимую модификацию белков-мишеней и тем самым регулировать их активность].
Сигнальный путь Wnt/β-катенин (Wnt wingless-type signaling pathway) – филогенетически древний механизм регуляции развития и поддержания гомеостаза тканей за счет контроля пролиферации (деления), дифференцировки стволовых клеток, миграции и апоптоза (запрограммированной смерти клеток). Активный участник процессов регенерации, роста костей, и массы других процессов, связанных с морфогенезом и определением клеточной судьбы. Катаболический путь для хрящевой ткани (для диска).
Сигнальный путь Ядерного фактора «каппа-би» [официально: транскрипционный фактор NF-κB; англ. nuclear factor kappa-light-chain-enhancer of activated B cells]; центральный компонент клеточного ответа на повреждение, стресс или воспаление. Мастер – регулятор катаболических и воспалительных реакций клеток: функционирует как один из важнейших быстродействующих внутриклеточных мессенджеров, соединяющий самые разнообразные сигналы окружающей среды с последующей экспрессией многочисленных генов. NF-κB оперативно регулирует различные биологические процессы, начиная от роста клеток, их выживания до развития тканей и органов. Играет ведущую роль в патогенезе многих хронических воспалительных процессов. NF-κB регулирует многие гены, участвующие в кодировании цитокинов, хемокинов, белков острой фазы, молекул адгезии, индуцибельных эффекторов ферментов (например, циклооксигеназы-2 (ЦОГ- 2; СОХ-2), таким образом являясь активатором иммунных и воспалительных реакций. NF-κB является центральным звеном активации генов, кодирующих регуляторы апоптоза и клеточной пролиферации.
Применительно к болям в спине: активация NF-κB повышает синтез матриксных металлопротеиназ (ММР -1, 3, 13) и аггреканаз (ADAMTS 4,5) в хондроцитах хрящей суставов и хондроцитоподобных клетках диска в ответ на механический стимул (компрессия), тканевую гипоксию, оксидативный стресс, продукты гликации, клеточный детрит («мусор»), ацидоз (снижение pH диска).
Матриксные металлопротеиназы (Matrix metalloproteinases; ММР) – основные протеолитические энзимы в организме, разрушающие белковые структуры (матрикс тканей). Их синтез приводит к «расплавлению» матрикса диска или хрящевых поверхностей суставов.
Аггреканазы (A Disintegrin and Metalloproteinase with Thrombospondin motifs; ADAMTS) – основные протеолитические энзимы, разрушающие аггреканы, белковые комплексы, удерживающие воду в тканях.
Сигнальный путь Митоген – активируемой протеинкиназы (MAPK) – нечто среднее между NF-κB и Wnt/β-катенин сигналингом: клеточный ответ будет зависеть от стимула.
В «семью» MAPK входят ERK, p38MAPK и JNK. Экстраклеточный-сигнал регулируемая киназа (ERK) активируется митогенами, ростовыми факторами, трансформирующими факторами роста, факторами роста фибробластов и инсулиноподобным фактором роста, таким образом регулируя рост клеток, их дифференциацию, развитие и жизненный цикл. Сигналинг ERK поддерживает выживаемость клеток при гипоксии и перепадах осмотического давления. Механочувствительный (активируется интегринами). При активации провоспалительными цитокинами фактором некроза опухоли-альфа (ФНО- α; TNF-α) и интерлейкином -1 бета (ИЛ-1β; IL-1β) поддерживает воспаление.
p38MAPK – стресс-активируемый путь, регулирующий воспаление, апоптоз, дифференциацию клеток. Активируется гипоксией, гиперосмолярностью, механической нагрузкой, эндоплазматическим стрессом, ацидозом, высоким уровнем глюкозы и провоспалительными цитокинами.
JNK (янускиназный) путь чем-то схож с p38MAPK, только он «легок на подъем» при активации пропионовыми бактериями и легко запускает апоптоз. Активация JNK приводит к повреждению ДНК, повышению синтеза матриксных металлопротеиназ, активации Toll – подобных рецепторов 2 типа (ниже в тексте), индукции синтеза «белка старения» р53. Принимает активное участие в дегенерации дисков.
Суммарно NF-κB и MAPK пути определяют активацию генов, которые регулируют синтез матрикс деградирующих энзимов (ADAMTS 4,5, ММР – 1, 2,3,9, 13, 14) и их антагонистов – тканевых ингибиторов матриксных протеиназ (TIMP-1, 2); матриксных протеинов (аггрекана, коллагена I и II типа), регулируют синтез простагландинов (через ЦОГ – 2); цитокинов ИЛ-6 (IL-6), ИЛ – 8 (IL-8) ; ростовых факторов IGF-1 и TGF – beta1; транскрипционного фактора SOX-9 (влияет на Wnt сигналинг хондроцитов); синтез оксида азота и многое другое.
При этом активация ядерного фактора «каппа би» (NF-κB) помимо выработки провоспалительных цитокинов (IL-1β, TNF-α, IL-6 и др.), в малых количествах вызывает преждевременное «старение» клеток со сменой их фенотипа, в большом количестве – приводит к смерти клеток (апоптозу – «запрограммированной», но часто преждевременной, или к пироптозу – преждевременной смерти вследствие критического повреждения самой клетки). В любом случае, гиперактивация данного транскрипционного фактора не сулит ничего хорошего ни для самой клетки, ни для окружающего ее матрикса, ни для ткани, ни для органа.
Нарушение регулировки сигнальных механизмов, основой которых является NF-κB и MAPK сигналинг, связано со многими тяжелыми болезнями человека, такими как рак, аутоиммунные заболевания, хронические воспаления, нарушение обмена веществ, диабет обоих типов, нейродегенеративные болезни, ревматоидный артрит, остеоартриты, дегенеративная болезнь диска и прочее.
Hedgehog («ежовый») сигналинг – критическая роль в формировании костей: отвечает за дифференцировку остеобластов, хондрогенез, формирование хрящей. Hedgehog (тип Sonic) стимулирует формирование остеокластов и препятствует сращению костей. Участвует в прогрессировании возраст-зависимых заболеваний (сердечно-сосудистые заболевания, старение кожи).
FOX0 (Forkhead box) – катаболический путь для мышц и хрящевой ткани.
NOTCH – регулирует гомеостаз костей. «Ишемический» (зависящий от кровоснабжения) сигнальный путь, завязанный на парциальном напряжении кислорода. Подавляет апоптоз в клетках пульпозного ядра и фиброзного кольца после активации их фактором некроза опухоли-альфа или при гипоксии. Сигнальный путь NOTCH пересекается с NF-κB, MAPK, PI3/AKT и Wnt/β-катениновым путями, влияя на их сигналинг.
Rhoa/ROCK (Ras homolog family member A/Rho-associated protein kinase 1 (ROCK)) – механический путь, активируемый интегринами. Пересекается с Wnt/β-катенин сигнальным путем. Регулирует синтез генов L-Sox-5, Sox-6 и Sox -9, от которых зависит синтез коллагенов I и II типов – основного материала матрикса связочной и хрящевой ткани.
TGF/BMP (путь трансформирующих ростовых факторов/костных морфогенных протеинов) – регуляция роста кости через контроль дифференцировки остеобластов. BMP регулируется миокином Ирисином через интегрины. Снижение BMP сигналинга связано с возрастным остеопорозом.
FGF (фактор роста фибробластов) сигналинг – развитие скелета и дифференцировка фибробластов: стимуляция пролиферации предшественников остеобластов (самообновление костей), регулирует гомеостаз фосфатов, метаболизм витамина D, пересекается с сигнальными путями Wnt и p53/p21(сигнальный путь старения клеток), которые вовлечены в патофизиологию остеопороза и кальцификации сосудов. Взаимодействует с сигнальными путями MAPK и PI3/AKT, опосредовано изменяя их сигналинг.
Сигнальный путь PI3/AKT/mTORC1. (Фосфоинозитид-3 киназа/протеинкиназа В/механическая цель рапамицина). На данном сигнальном пути держится вся физическая реабилитация. В зависимости от типа сигнала данный путь активирует или анаболические гены, или катаболические. Данный путь пересекается с NF-κB и MAPK путями и напрямую подавляет их активность при условии анаболической стимуляции.
PI3/AKT – анаболический путь для хрящевой и связочной ткани. mTORC1 – мастер-регулятор мышечного анаболизма. Если сигнальный путь Wnt/β-катенин специализируется на метаболизме костей, то PI3/AKT/mTORC1 – это метаболизм мягких тканей опорно-двигательной системы.
Существует множество других сигнальных путей, клеточных мессенджеров и транскрипционных факторов активации генов с последующим развитием внутриклеточных, экстраклеточных и тканевых/внутриорганных событий.
Многие из сигнальных путей перекрываются и «переплетаются» между собой, зачастую используя общие сигнальные молекулы/мессенджеры (цитозольные медиаторы). Грубо говоря, сигнальные пути можно сравнить с химическими «дорогами» с их перекрестками и развилками.
Мембранные рецепторы взаимодействуют друг с другом через адапторные белки, при этом сигналы с рецепторов распространяются по различным путям, одновременно активируя различные типы сигнальных мессенджеров/посредников, создавая перекрестные помехи между внутриклеточными сигнальными каскадами. Механические и химические механизмы передачи сигнала стимулируют одни и те же внутриклеточные сигнальные пути (ни один из них не работает изолированно от другого), при этом вызывая синергетические и конкурирующие эффекты, направленные либо на усиление, либо на подавление генетического ответа.
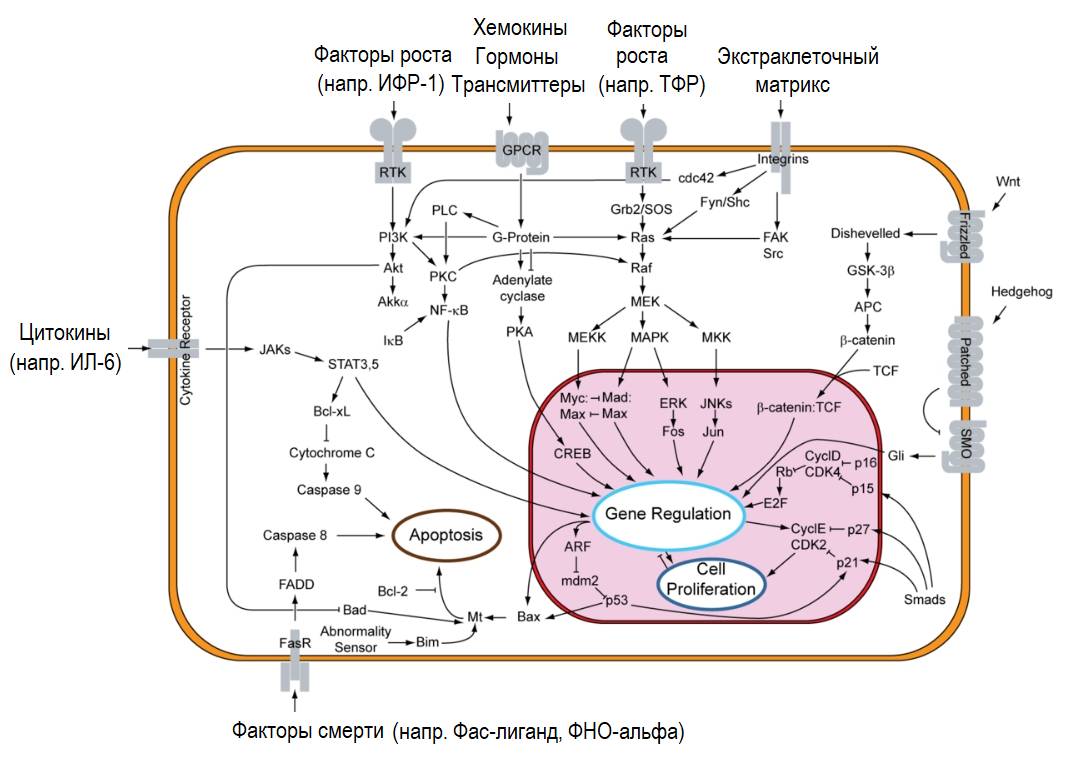
Общий клеточный сигналинг (Википедия).
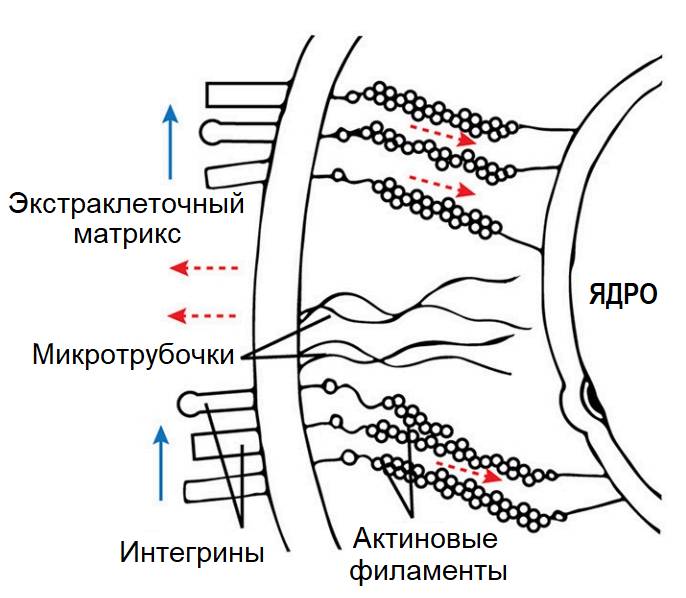
Актиновые «вожжи».
Механический сигнал через интегрины может передаваться напрямую через систему белковых нитей, соединяющих экстраклеточный матрикс с мембраной клетки, и нитей «цитоскелета», пронизывающих внутриклеточный цитозоль и ведущих напрямую к оболочке ядра. Задействован принцип «вожжей». Это дает выигрыш в скорости биохимической реакции клетки на внешние стимулы. Таким образом, механические нагрузки влияют на транскрипцию генов опосредованно через активируемые рецепторами сигнальные каскады и напрямую через изменение транспорта ядерной мембраны. Поэтому клетку можно представить как компьютер, который суммирует все внешние механические, химические и электрические сигналы и вычисляет, какой ответ в итоге будет выдан в «сборочный цех» клетки: анаболический или катаболический. Синтезированные в результате реакции клетки функциональные белки являются основными факторами в текущем поведении клетки (интракринное/аутокринное влияние) и определяют ее дальнейшую судьбу. Клетка может ответить синтезом структурных белков, используемых для ремоделирования матрикса, или выдать провоспалительный ответ в виде синтеза цитокинов и протеиназ, разрушающих матрикс, или сменить фенотип (мгновенно «постареть»), или активировать режим самоуничтожения (запланированный – апоптоз, или вынужденный вследствие повреждения мембраны – пироптоз и т.п.).
Таким образом, дифференцировка, рост, жизненный цикл и гибель клеток регулируются и всецело зависят от внешних стимулов и внутриклеточных сигнальных каскадов, влияющих на генетическую регуляцию.
Механические силы, такие как растяжение, сжатие и сдвиг, воспринимаются и передаются на границах раздела «клетка-клетка», «клетка-матрикс» и «клетка-межклеточный просвет», создавая реакции, которые могут либо усилить структуры, подвергающиеся механическому воздействию, либо изменить фенотип или структуру клеток и в дальнейшем привести к дистрофии или деградации/дегенерации тканей вследствие гибели клеток.
Посредством молекул клеточной адгезии (слипания) клетки также оказывают прямое механическое влияние на соседние клетки. Эти молекулы включают трансмембранные белки, такие как интегрины, кадгерины, селектины и коннексины, которые связывают клетки вместе для структурных и функциональных целей. Соединения между клетками очень напоминают клеточные матричные комплексы с динамическими связями с цитоскелетом, которые реагируют на механическую нагрузку. Эти контакты влияют на «коллективное» поведение клеток, включая адгезию, миграцию, изменение формы, пролиферацию, дифференцировку стволовых клеток, передачу внутриклеточных сигналов и обмен матрикса.

