


Полная версия:
Психотроника и днк. Искусственный интеллект в битве со старением организма человека. Психотроника и днк
С другой стороны, очень сложные для дискретного вычисления двумерные преобразования Фурье предельно просто (одной сферической поверхностью) выполняются в оптической системе, использующей когерентные волны, а свойства таких систем можно увидеть в ядрах мозга. Кора мозга, четко проявляет свойства голографической памяти. Пучки волокон, неискаженно переносящие образы от ядер к коре и от коры к ядрам, замыкают цепочку совпадений между ожидаемой вычислительной структурой и реальностью. Так мы снова приходим к нейрокорреляторам. Эксперименты показали, что «алгоритм обработки зашумленных изображений в зрительной системе наблюдателя можно моделировать алгоритмом, в соответствии с которым работает приемник Зигерта – Котельникова, дополнив его „логическим ограничением уровня яркости“ на выходе этого приемника» [Красильников, тысяча девятьсот восемьдесят шестой год, страница сто шестая]. Теперь становятся понятными вечные проигрыши электронных распознающих устройств в соревновании с человеком. Ни в одной из практических распознающих систем, в разработке которых участвовал или с которыми знакомился автор, голографические корреляторы не применялись. Дело в том, что голографический коррелятор, использующий лучи видимой части спектра, является очень деликатным прибором, которое трудно перенести из лаборатории в практику, а его электронно-вычислительные имитации работали медленно. Поэтому реальные устройства распознавания зрительных образов строились по иным схемам. Отгого-то они и были обречены на постоянный проигрыш перед человеком, в мозге которого реализована теоретически наилучшая, предельная по возможностям схема приемника Зигерта – Котельникова.
Адресация в коре мозга
В технических системах голографической памяти изменение адреса записываемого или считываемого изображения, т.е. поворот опорного (или считывающего) пучка лучей относительно запоминающей среды, выполняется путем механического поворота фотопластинки, зеркала или другого элемента оптической схемы. Мозг, как известно, не содержит движущихся частей. Природа выбрала иной вариант управления направленностью волн, не требующий механических перемещений, а потому более быстрый и надежный. Она использовала хорошо известный в радиотехнике (особенно – в радиолокации) принцип фазированной антенной решетки. Его суть в следующем. Если расположить в ряд несколько антенн (например, ультра коротких волн – диполей) и подать на них одинаковый, строго фазированный сигнал, то волна излучения будет распространяться в пространстве перпендикулярно этому ряду. Если же изменить фазы подаваемых на диполи колебаний так, чтобы фаза сигналов антенны с более высоким номером отставала на определенную величину от фазы предыдущей антенны, то суммарный вектор излучения (вектор Пойнтинга) антенного ряда отклонится от перпендикуляра в направлении более высоких номеров антенн и наоборот. Угол отклонения растет с ростом разности фаз между соседними антеннами. При необходимости управления вектором Пойнтинга в двух взаимно перпендикулярных направлениях разработчики используют параллельно расположенные ряды антенн, т.е. двумерную антенную решетку.
Чтобы управлять величиной фазового сдвига между элементарными антеннами, разработаны устройства, называемые фазовыми вращателями. Нервные окончания пучка волокон, приносящего волну-образ, например, к одному из участков коры больших полушарий, могут рассматриваться как элементарные излучатели (антенны) фазированной антенной решетки. Если немодулированный фронт опорной волны нервного возбуждения достигнет всех окончаний одновременно, в одной фазе, то общий вектор излучения окажется перпендикулярным поверхности решетки. Если же волна возбуждения появится у одного края решетки раньше, чем у другого, то вектор распространения возбуждения в тканях коры отклонится от перпендикуляра в сторону второго края решетки.
Многие нейробиологи полагают, что скорость распространения нервного возбуждения в одной и той же нервной ткани неизменна, как неизменно время прохождения нервного импульса по аксону. Но это не так. Даже в отдельном нейроне под влиянием тормозных воздействий может несколько изменяться латентный период между поступлением внешнего стимула и возникновением потенциала действия. Главное же в другом – нервная ткань ведет себя не так, как отдельный нейрон.
В зависимости от соотношения тормозных и возбуждающих воздействий, нейрон может возбудиться при подходе волны возбуждения или «промолчать». Но молчание части нейронов вовсе не прекращает распространение волны возбуждения по ткани. Разница лишь в том, что в зоне торможения нервное возбуждение распространится не кратчайшими путями, а по лабиринту более редкой сети нейронов, сохранивших активность, на что потребуется больше времени.
Это значит, что тормозное воздействие способно снизить среднюю скорость распространения возбуждения по нервной ткани. Поэтому мощное тормозящее воздействие вблизи зоны окончаний нервного пучка на поверхности коры способно, по принципу фазированной антенной решетки, отклонить в свою сторону вектор перемещения энергии (волны) нервного возбуждения – вектор Пойнтинга.
На рисунке показана схема поворота (в одной плоскости) вектора излучения волны нервного возбуждения в коре мозга при изменении адреса записываемого или считываемого образа.
Здесь показаны: 1 – пучок нервных волокон, подводящий к коре мозга немодулированную опорную волну нервного возбуждения при запоминании, или же считывающую волну – при считывании образа из памяти; 2 – пучок нервных волокон, подводящий волну-образ с сенсорной информацией («предметную» волну – в терминах теории голографических систем); 3 и 4 – сигналы тормозных воздействий, управляющие адресацией; 5- область градиентного торможения, изменяющая направление волн нервного возбуждения в коре мозга; 6 – вектор Пойнтинга одной из опорных (или считывающих) волн; 7 – фронт одной из опорных (или считывающих) волн; 8 – фронт волны-образа с сенсорной информацией при записи; 9 – нервная ткань коры мозга, являющаяся голографической запоминающей средой: 10 – пучок нервных волокон, отводящий при считывании волну нервного возбуждения с извлеченной информацией.
Представим себе, что в точке, куда подходит сигнал 3, приложено мощное тормозное воздействие, распространяющееся в области 5 и плавно уменьшающееся по мере удаления от точки приложения. Это приведет к неравномерному уменьшению скорости распространения волн нервного возбуждения при прохождении области 5 – снижение скорости окажется наиболее заметным вблизи тормозного воздействия и все менее ощутимым с удалением от него. Соответственно, вектор Пойнтинга и фронт опорной (или считывающей) волны окажутся повернутыми в сторону тормозного воздействия, что определит адрес голографической записи. Чем сильнее торможение – тем на больший угол отклонятся вектор и фронт волны.
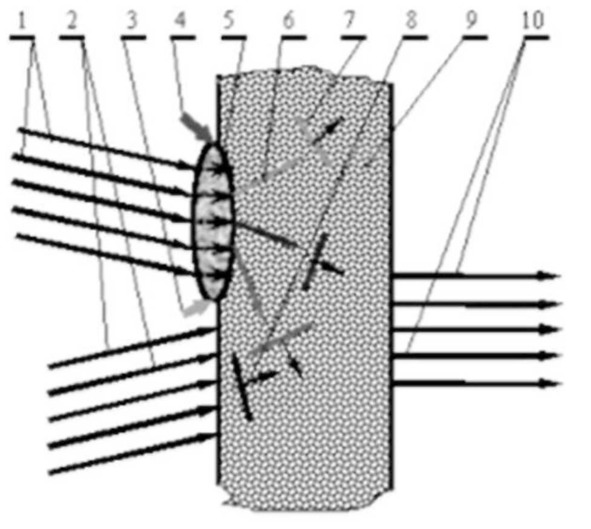
Схема поворота вектора излучения волны нервного возбуждения в коре мозга при изменении адреса записи-считывания.
The scheme of turn of a vector of radiation of a wave of nervous excitation In a bark of a brain at change of the address of record-reading.
Схема, изображенная на рисунке, показывает устройство системы адресации в одной плоскости. Но аналогичная картина имеет место и в перпендикулярном направлении, что обеспечивает повороты вектора Пойнтинга опорной и считывающей волн, ориентировочно, в пределах полусферы. При записи нового образа опорная и предметная волны нервного возбуждения интерферируют между собой, создавая в коре мозга сложную картину распределения возбужденных областей. Запоминается именно пространственное распределение возбужденных точек интерференционного поля. Если в дальнейшем считывающая волна пройдет под таким же углом, как и опорная волна при записи, то окажется воспроизведенным образ, записанный по данному адресу. Изменение величин тормозящих воздействий или их пространственного распределения изменят адрес записи-считывания. На рисунке для удобства изображения схемы отведение извлеченного из памяти образа показано с противоположной (относительно подводящего пучка) стороны коры. Реальное считывание происходит с той же стороны, что и запись.
Таким образом, в момент запоминания новых данных в коре мозга одновременно распространяются и пересекаются (интерферируют) две волны – волна-образ, несущая сенсорную информацию, и немодулированная опорная волна. Последняя не несет никакой информации, но без нее процесс голографической записи в нервной ткани был бы невозможен. Изменение интенсивности тормозных процессов в зоне подведения опорной волны к коре мозга регулирует направление распространения каждой очередной волны и определяет адреса записи новых образов в нейроголографической системе.
В процессе восприятия или осмысливания сенсорной информации, при определении корреляции нового образа с данными, ранее записанными в памяти, происходит одновременное сопоставление со многими образами, формируется двумерное поле результатов сравнения, содержащее многочисленные пики корреляционной функции. Важнейшим является пик максимальной высоты. Он указывает адрес образа, наиболее сходного с анализируемым.
Для извлечения из памяти (вспоминания) конкретного образа нужно направить немодулированную считывающую волну точно по его адресу или, иначе говоря, точно под таким углом, под каким проходила опорная волна при записи. За жизнь человека мозг запечатлевает огромное количество волн-образов и, если мы восторгаемся меткостью стрелка, попадающего в центр мишени, то не меньшего восхищения заслуживает человек с хорошей памятью, ибо его мозг с точностью снайпера направляет считывающие волны за нужной информацией строго по нужным адресам. К сожалению, такая точность работы мозга реализуется далеко не у всех людей.
Мысль-организованная группа волн
Представление о прохождении волны-образа через нейрокоррелятор близко к известному выражению «мелькнула мысль». Но элементарная осознаваемая мысль не эквивалентна прохождению единственной волны- образа. Элементарный акт мышления включает в себя обособленную, но внутренне функционально связанную группу волн нервного возбуждения, в которой каждая волна выполняет особую, индивидуальную функцию.
Данные исследований позволяют заключить, что мысль, как правило, включает в себя от трех до десяти волн нервного возбуждения, причем запоминается и потому осознается только исходная информация и общий результат этого составного процесса.
Например, в одной из серий экспериментов [Sheatz, Chapman, тысяча девятьсот шестьдесят девятый год] регистрировались волны электроэнцефалограмм при исследовании отношения человека к различным стимулам. Вызванные потенциалы состояли как минимум из трех последовательных волн. Если испытуемый наперед знал, что сигнал не имеет значения и не требует ответа, то амплитуды второй и третьей волн снижались. Если он должен был сначала правильно определить сигнал, а затем мог не отвечать и больше не обращать на него внимания – тогда уменьшалась только третья волна. И все три волны достигали максимума, когда испытуемому приходилось сперва на мгновенье задуматься над стимулом, а затем ответить на него. В подобных экспериментах особенно проявилась независимость управления каждой солитонной волной, что принципиально отличает их от синусоидальных колебаний.
Различие в функциях разных волн цикла, вероятно, вытекает из различия задач, решаемых каждой из них. Во-первых, пришедшая информация должна быть зафиксирована в голографической памяти, что требует прихода одновременно с волной-образом и немодулированной опорной волны. Во- вторых, новый образ должен быть классифицирован, должен пройти сравнение с несколькими из хранящихся в памяти эталонов, для чего может понадобиться несколько волн нервного возбуждения. В-третьих, после отыскания наилучшей корреляции результат должен быть подан на выход, и зафиксирован в последующих отделах памяти, где регистрируются уже не образы, получаемые извне, а итог их корреляционного сравнения с ранее записанными, известными образами. В промежутке между прохождением двух солитонов, в нервной ткани, не относящейся к области памяти, происходит «стирание» предыдущей информации, чтобы она не повлияла на независимое протекание дальнейших вычислений. Обычно такое «стирание» имеет вид перехода ткани к хаотической импульсной активности. По-видимому, характер этих процессов сходен с характером процессов в зрительном анализаторе. Вот как описывается цикличность работы нашего зрения в связи с саккадическими «скачками» глаза. «В первый момент после скачка… система имеет характеристику фильтра низких пространственных частот,… способна различать только крупные детали изображения… В следующие моменты разрешающая способность увеличивается… Процесс идет непрерывно от пятидесяти до семидесяти мили секунд и к концу этого времени… имеет… максимальную разрешающую способность. В этом оптимальном состоянии система находится от тридцати до пятидесяти мили секунд … /Затем/ … происходит… функциональная дезорганизация полей.
Благодаря этому стирается «предыстория» … и они становятся готовыми к новому циклу восприятия…» [Подвигин, тысяча девятьсот семьдесят девятый год]
Прямое приложение принципов КНГ к зрительному анализатору человека обнаруживает два резких несоответствия между свойствами голографических корреляторов и свойствами нашего зрения. Во-первых, корреляторы инвариантны (безразличны) к плоскопараллельному смещению анализируемого объекта в поле зрения, а человек к этому вовсе не безразличен, он упорно сосредотачивает взгляд на рассматриваемом предмете. Во-вторых, человек без малейшего напряжения узнает приблизившийся объект, если узнал его издали, тогда как для коррелятора это невозможно. Коррелятор работает только при том масштабе изображения, какой заложен в эталоне. Анализ показал, что объяснение этих противоречий заключено в особой анатомии глаза и зрительного нерва. Система нейронов зрительного анализатора человека построена так, что наблюдаемое изображение деформируется путем сжатия его периферийных областей. Сжатие, насколько можно судить, соответствует логарифму расстояния от центра поля зрения. Деформирование изображения происходит под совместным действием трех факторов:
от центра к периферии сетчатки глаза уменьшается плотность расположения фоторецепторов;
– от центра к периферии сетчатки растет число рецепторов, посылающих сигналы в одно и то же волокно зрительного нерва;
на входе зрительного нерва в ядро, называемое наружным коленчатым телом таламуса, наблюдается редкое явление – плотность расположения нервных окончаний оказывается наиболее высокой на периферии пучка волокон и плавно уменьшается к центру.
Реализуемая таким способом логарифмическая деформация входного изображения превращает первый голографический коррелятор зрительной системы в одну из его разновидностей, в так называемый коррелятор Меллина [Василенко, Цыбулькин, тысяча девятьсот восемьдесят пятый год]. Отличительной особенностью коррелятора Меллина является инвариантность к размерам анализируемого образа. Иначе говоря, коррелятор Меллина, как и наше зрение, способен распознавать знакомые объекты при различной угловой величине, т.е. на разном расстоянии, лишь бы они были хорошо различимы. Это достигается нелинейностью (логарифмическим масштабом) системы относительно центра поля зрения, что влечет за собой необходимость центрирования распознаваемого образа. Следовательно, свойства нашей зрительной системы, как распознающего устройства, точно соответствуют специфике того типа голографического коррелятора, к которому она должна быть отнесена на основании ее анатомических характеристик. Кстати, это лишний раз подтверждает голографическую природу вычислений.
О мышлении в целом
Данные, говорящие о существовании в мозге, по крайней мере, одного нейронного коррелятора. Но что говорит о существовании пирамидальной иерархической структуры из многих корреляторов? Подтверждение этому дало интересное общее свойство мозга, обнаруженное при исследовании условных рефлексов. Эксперименты выявили некую универсальную закономерность, понять природу которой исследователям не удалось.
Закономерность, о которой идет речь, обнаружилась при изучении вариаций биопотенциалов мозга подопытного животного в ответ на изменения воздействий внешней среды. После того, как рефлекс выработан, кривая активации биопотенциалов в ответ на любое изменение внешних условий приобретала сходство с кривой частотного резонанса, как показано на рисунке.
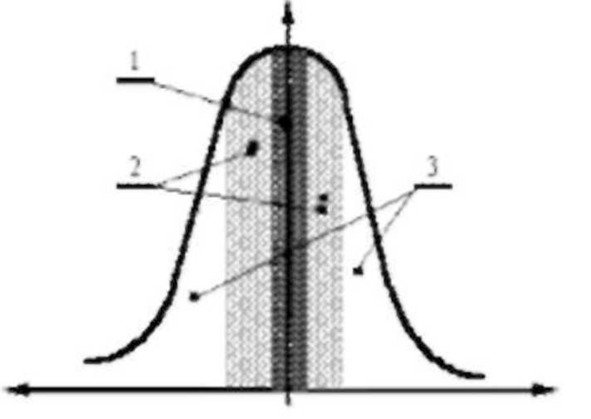
Рисунок Зависимость активации биопотенциалов мозга от изменения параметров сигнала при выработанном условном рефлексе [Кратин и др., тысяча девятьсот восемьдесят второй].
1 – область специфических условно рефлекторных реакций;
2- область не специфических вегетативных эффекторных ответов;
3 – область электроэнцефалографического проявления активации биопотенциалов мозга без эффекторных ответов.
Figure Dependence of activation of biopotentials of a brain on change of parameters of a signal At the developed conditioned reflex [Kratin, etc., one thousand nine hundred eighty second].
– area of specific conditionally reflex reactions;
area nonspecific vegetative effektornyh answers;
– area elektroentsefalograficheskogo displays of activation of biopotentials of a brain without effektornyh answers.
Однако, вопреки стараниям исследователей, самое настойчивое экспериментальное изучение всех возможных «кандидатов» не выявило ни в мозге, ни в организме в целом, колебательного процесса, на который можно было бы возложить ответственность за такой «резонанс» [Кратин и др., тысяча девятьсот восемьдесят второй год; Кратин, тысяча девятсот восемдесят шестой].
Другая удивительная особенность явления состояла в том, что достижение пика электрической активности мозга зависело не от одной какой-то характеристики, одного раздражителя, а от целостного комплекса параметров, от всей суммы условий, существовавших во время исследований. Достаточно изменить любой параметр эксперимента или обстановки, окружавшей животное в ходе выработки условного рефлекса, чтобы изменился уровень активации биопотенциала.
То, что осталось загадкой для исследователей, получило четкое объяснение с позиций КНГ. Перед учеными оказалась не резонансная кривая, а очень похожая на нее кривая функции корреляции информационных массивов.
Тот факт, что кривая во всех случаях отображает зависимость активации биопотенциалов мозга от целостного сенсорного образа внешнего мира, говорит как раз об объединении сигналов разных сенсорных систем по мере подъема по пирамиде принятия решений. Говорит о таком объединении сигналов в пирамиде корреляторов, которое, в конце концов, приводит к Формированию единого сенсорного образа на входе нейронного коррелятора высшего уровня.
Почему я отстаиваю и развиваю именно концепцию нейроголографии? Этот принципиальный вопрос требует четкого ответа.
а) Многократная нехватка памяти ДНК для описания мозга как ассоциативной сети показала, что геном описывает, в основном, не клеточный, а более высокий уровень организации мозга – уровень больших ансамблей нейронов, слоев коры, пучков волокон, скоплений нейронов в виде ядер и т. п. Но из таких элементов, как слои, пучки и ядра можно создать лишь один тип вычислительных и, вообще, информационных систем
только оптические информационные системы.
б) На организацию мозга по типу оптической информационной системы указало и то, что вместе с ним Природа создала в ходе эволюции два типа нервных тканей, специально приспособленных для обеспечения оптических закономерностей. Это, во-первых, серое вещество мозга, отличающееся малыми размерами нейронов, большим количеством синапсов и диффузным распространением волн нервного возбуждения, в соответствии с принципом Гюйгенса. Во-вторых, это пучки нервных волокон, перенос нервного возбуждения по которым, в информационном смысле, аналогичен переносу оптического изображения по стекловолоконным жгутам.
в) Пучки нервных волокон, топологически точно переносящие информационные массивы со своих входов на выходы, получили, с появлением мозга, широкое распространение вопреки тенденции к повышению хаотичности нервной системы. Это доказывает особую роль в работе мозга топологически организованных двумерных информационных массивов, т.е. изображений или образов.
г) Низкий уровень фоновой активности нервных тканей мозга (5—7%) подтвердил, что регистрируемые энцефалографами волны интегральной импульсации нейронов имеют форму солитонов. А это, в свою очередь, объяснило их способность эффективно переносить двумерные информационные массивы в виде образов, модулирующих передний фронт пространственной волны. Синусоидальные волны не удалось бы модулировать подобным образом.
д) Внимание исследователей давно привлекло совпадение необычных свойств памяти мозга и голографической памяти – раздельная запись разных информационных массивов в общем пространстве памяти; распределенная запись, мало чувствительная к шумам и локальным повреждениям; быстрый вызов любой информации, независимо от адреса; огромная скорость записи и считывания при медленных элементарных процессах, что говорит о записи за одну операцию целостных образов, а не последовательности данных.
е) На эквивалентность мозга голографическому коррелятору указало полное соответствие человека свойствам оптимального приемника, т.е. свойствам теоретически наилучшей распознающей системы, каким является голографический коррелятор при восприятии двумерных образов в шумах.
ж) Организация мозга в виде системы голографических корреляторов подтверждается также анатомическими данными и точным совпадением с их свойствами не только сильных, но и слабых сторон вычислительной системы мозга. Известно, что мозг предельно легко распознает образы – даже ребенок узнает обстановку и людей, что относится к очень сложным задачам. Мозг быстро оценивает корреляцию между массивами данных, хорошо решает задачи в условиях неопределенности и шумов, что характерно для голографического коррелятора. Но даже искушенный в расчетах инженер с трудом решает в уме более простую задачу перемножения многозначных чисел, а это уже совпадает с трудностями выполнения арифметических операций голографическими корреляторами.
з) Расположение фоторецепторов глаза млекопитающего и особенность их коммутации, а также анатомия нервного пучка на входе наружного коленчатого тела таламуса, выявили деформацию зрительного образа, близкую к логарифмической, что характерно именно для одной из разновидностей голографического коррелятора – для коррелятора Меллина. Специфика данного коррелятора (инвариантность к масштабу и отсутствие инвариантности к сдвигу) демонстрируется и зрительной системой человека.
и) Характер зависимости биопотенциалов мозга от изменений стимула при выработанном условном рефлексе подтвердил организацию системы принятия решений в виде пирамиды нейрокорреляторов, а также поэтапное объединение сигналов различных сенсоров по мере перехода на верхние уровни пирамиды, что точно соответствует концепции нейроголографии в целом.
Отметим финальную часть про наших физиков. Взгляды физиков двадцатого столетия формировались на базе сложнейших экспериментов, в которых использовались, например, ускорители частиц, для сооружения которых приходилось объединять усилия ряда стран, а мировоззрение мистиков сложилось тысячелетия назад, без всяких приборов, в процессе медитации, напрочь отключающей в мозгу аппарат логического мышления.
Выявленный феномен бросает вызов современной науке и, прежде всего, физиологии мышления. Почему отключение логического мышления помогает проникновению в глубочайшие таинства материи и Вселенной? Каким образом медитация может дать сведения об удивительных и тяжело познаваемых законах квантовой механики, теории относительности, о других физических откровениях? Нельзя забывать, что мозг является информационно-вычислительным устройством квантово-механического уровня. Для возбуждения рецепторной клетки глаза достаточна одна субатомная частица – фотон. По нервному волокну сигнал возбуждения передается от одного перехвата Ранвье к другому под действием калий- натриевых насосов, в итоге переносящих немногочисленные субатомные частицы – электроны. При передаче сигнала через синоптическую щель та же задача решается единичными молекулами нейромедиатора. Вся динамика работы мозга основана на тонких операциях с субатомными частицами, отчего главными законами мироздания для мозга оказываются законы квантового мира. Это объясняет все стороны феномена, замеченного и описанного Фритьофом Капрой. Становится понятно, почему в отрешенном состоянии человек ощущает себя слившимся со всей Вселенной. Это не ошибка, не иллюзия, он, действительно, слит с представившейся внутреннему взору Вселенной, потому что эта Вселенная – его собственный мозг.

