
Полная версия:
Палеонтология антрополога. Три эры под одной обложкой
Среди головоногих в девоне неплохо себя чувствовали наутилоидеи Actinocerida, Orthocerida и Oncocerida, от последних тогда же возникли Nautilida. Раковина онкоцератид была загнута как рог, иногда даже на полтора оборота, причём завитки не смыкались. У первых наутилид она ещё была такой же, но в последующем раковина расширялась, завитки смыкались, пока не превратились в то лаконичное совершенство, что мы видим у современных наутилусов.
Головоногие Bactritida хотя сами по себе не были особо распространены и разнообразны, дали в самом начале девона начало важнейшей и богатейшей группе Ammonoidea, ставшей едва ли не символом палеонтологии. В этой эволюционной линии исходно и независимо произошло загибание прямой как стрела раковины. Например, у продвинутого бактритида Lobobactrites – сначала в бычий рог, потом в бараний, а потом и в плотную спираль. Раковина древнейшего раннедевонского аммонита Anetoceras была промежуточной: на ранних стадиях развития она формировалась как слабоизогнутая, а потом заворачивалась гораздо плотнее, отчего по итогу получалось колесо с отверстием в центре, в котором каждый следующий оборот почти, но не совсем – с узеньким зазором – смыкался с предыдущим. У чуть более поздних аммонитов раковина обычно просто свёрнута в плотную спираль. Забавно, что эмбриональные раковины девонских аммоноидей показывают в ходе индивидуального развития отличный переход от слабоизогнутой формы к согнутой и далее спирально закрученной; это один из отличных примеров биогенетического закона Геккеля. Поскольку раковина нарастала от крошечных внутренних колечек к огромным наружным не только в толщину, но и в ширину, поперечное сечение приобретало в итоге вид песочных часов: центральные завиточки совсем узенькие, а крайние – очень широкие. Одновременно такая форма повышала устойчивость при плавании, а у продвинутых видов стабильность дополнялась продольным килем. Внутри раковина аммонитов подразделена на отдельные камеры, которые моллюск заполнял через жабры газом или, наоборот, закачивал туда воду, меняя тем самым плавучесть. Вряд ли такая конструкция могла очень быстро двигаться по горизонтали, зато оперативно погружалась или всплывала.
Встречались и оригинальные варианты. У Soliclymenia paradoxa, Kamptоclymenia trivaricata и Parawocklumeria paradoxa из фаменского века, то есть самого конца девона, раковина была треугольной! При этом у соликлимении раковина закручивалась на много оборотов; у камптоклимении на небольшое число, причём углы были отделены друг от друга бороздками; а у паравоклюмерии оборотов по сути вообще не было, а каждый угол представлял собой обособленный пузырь.
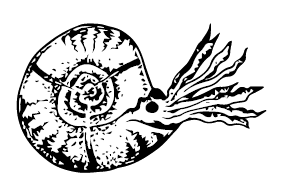
Kamptоclymenia trivaricata
После своего появления в начале девона аммониты дали гигантское количество видов – описано порядка трёх тысяч, но нет сомнения, что будут найдены и другие. Вплоть до своего вымирания в конце мела аммониты были самыми обычными жителями всех морей планеты. Такой успех стал возможен в немалой степени благодаря изобретению челюстей, что позволило аммонитам стать не только фильтраторами, но и хищниками.
В раннем девоне доживал свой век последний аномалокарид Schinderhannes bartelsi из Германии. В отличие от своих эпичных кембрийских предков, шиндерханнес (названный так, кстати, по имени бандита XVIII века из области, где найден отпечаток) был мелким, всего десяток сантиметров, и крайне пучеглазым. Спереди он имел пару длинных тонких «плавников», сзади – пару коротеньких «плавничков», а заканчивалось тело вытянутым шипом. Видно, что аномалокариды пытались соответствовать веяниям времени, но рыбы давно и безнадёжно их обогнали. Успех группы был в прошлом, аномалокариды не выдержали конкуренции и вымерли.
Та же судьба ждала и маррелломорф – раннедевонские Mimetaster hexagonalis с панцирем в виде шестиконечной остролучевой звезды и длинными тонкими членистыми ножками и щетинками, а также Vachonisia rogeri с панцирем странного остроугольного фасона в форме супергеройского плаща были последними представителями этой загадочной группы членистоногих. Перед вымиранием маррелломорфы были довольно успешны: миметастеры в некоторых местонахождениях находятся на втором месте по числу отпечатков после трилобитов. Возможно, они даже жили группами.
Трилобиты в девоне не скучали. Особенно колоритен раннедевонский североамериканский Terataspis grandis – шипастая шестидесятисантиметровая плюха с растопыренными во все стороны отростками и пупырчатым шаром на голове, очень похожая на современную морскую звезду «терновый венец». Главные мекки любителей девонских трилобитов – Оклахома и Марокко, тут находятся, наверное, миллионы окаменелостей изумительной сохранности; главное – уметь их отпрепарировать. Особенно причудливый облик приобрёли трилобиты Odontopleuroidea – их панцири разрослись совершенно безумными рогами, часто к тому же пупырчатыми, растопыренными во все стороны, а иногда даже завитыми в спирали. Чудесны оклахомские, чешские и германские Dicranurus elegans и D. monstrosus, Kettneraspis williamsi и Ceratonurus krejcii, марроканские Ceratarges ziregensis, Cyphaspis boutscharafinense, Kayserops megaspina, Kolihapeltis rabatensis, Koneprusia dahmani и Quadrops flexuosa. Все они прекрасны, трудно выбрать рекордсмена! Понятно, что не только шипами знамениты девонские трилобиты. Марокканский Erbenochile issoumourensis уникален высокими вертикальными глазами-щитами, смотрящими в разные стороны. У Walliserops trifurcatus на «носу» торчал огромный – длиннее всего тела – рог-трезубец. У Lanceaspis hammondi нос был длинным и треугольным, как у осетра. Головной щит Harpes perradiatus огромный, широкий полукруглый, в виде копыта, но с длинными отростками позади, достающими до самого конца тела. Ясно, что существовали и более простые формы, например Bellacartwrightia whiteleyi, тоже, впрочем, красивые.
Как и раньше, некоторые трилобиты – например, Trimerocephalus chopini – были не прочь потусоваться и отправиться в путь плотной цепочкой, касаясь друг друга, многочисленные примеры чего найдены в Польше.
Работа над ошибками
Марокканские трилобиты – это просто праздник какой-то. Они всем нравятся, а потому местные жители научились добывать их на продажу туристам. Однако препарировать одну супершипастую окаменелость долго и трудно, а туристы бывают привередливы – даже чуток обломанные рожки им уже не нравятся. Потому умельцы наловчились лепить трилобитов из смолы или даже из перемолотой и заново склеенной местной породы, да так искусно, что даже специалисты не всегда с первого взгляда распознают подделку.
Мечехвосты в девоне приобрёли уже почти современный вид, например Neobelinurus rossicus и Kiaeria limuloides.
Работа над ошибками
Название «Kiaeria» не выглядит каким-то особо пафосным или модным, однако ж, было дано трём совершенно разным существам – силурийской ракушковой ракообразной остракоде, девонскому мечехвосту и современному мху.
Ракоскорпионы достигли своего наибольшего великолепия: раннедевонский германский Jaekelopterus rhenaniae вымахал до 2,3–2,6 м! Это абсолютный рекорд среди всех членистоногих за всю историю планеты. Своими растопыренными клешнями он тиранил первобытных рыб и всех, попадавшихся ему на пути. Другие ракоскорпионы могли быть меньше, но не менее впечатляющими: Mixopterus kiaeri достигал 20–70 см и обладал огромными, возможно, ядовитыми иглами на педипальпах и передних ходильных конечностях, а также длинным шипом на «хвосте». У небольшого таймырского Borchgrevinkium taimyrense голова занимала почти половину длины.

Mixopterus kiaeri
Некоторые мелкие девонские ракоскорпионы начали заселять пресные водоёмы, где жили моллюски и рыбы, служившие им пищей.
Продолжали осваивать сушу скорпионы. Из раннего девона Германии известен не самый древний, зато самый примитивный и первый наземный скорпион Palaeoscorpius devonicus. Вероятно, он уже дышал примитивными лёгкими, а не жабрами. В девоне скорпионы уже окончательно и бесповоротно вышли из воды. А ведь в это время суша всё ещё представляла собой пустыню, лишь по берегам теснились заросли псилофитов. Скорпионам пришлось нелегко – им пришлось выныривать из воды сразу на раскалённые камни. Зато они так круто приспособились к крайне неблагоприятным условиям, что и сейчас отлично чувствуют себя и в пустынях, и в джунглях. Адаптации скорпионов оказались идеальны, и потому эти животные практически не поменяли свой внешний облик и не могут быть никем вытеснены из своей экологической ниши – она уже надёжно занята, пролезть туда невозможно. Опоздавшим пришлось приспосабливаться к другим местообитаниям, а потому больше меняться.
Маленькая тонкость
Между прочим, это универсальное правило эволюции: первый в новой экологической нише больше никого туда не пускает и сам застывает в состоянии «живого ископаемого» – нам на радость, позволяя составить представление о далёком прошлом. Другие существа, осваивая позже возникшие, а потому менее выгодные условия, вынуждены меняться. Таким образом, предшествующие стадии эволюции, как правило, долго никуда не деваются (если их не сотрёт с лица Земли очередное великое вымирание), а новые постоянно добавляются. Они сами меняют мир, создают дополнительные экологические ниши, и эволюционная куча мала продолжает нагромождаться. Оттого разнообразие жизни со временем в среднем увеличивается.
Продолжали эволюционировать и пауки – отлично сохранился североамериканский Attercopus fimbriunguis с древностью 374–386 млн л. н., он уже умел выделять паутину, но не умел её плести, так как не имел прядильных органов (сначала их вроде бы нашли, но на новом материале стало видно, что их таки нет). В девоне появляются наземные почвенные акариформные клещи Acariformes, например, Archaeacarus dubinini, минимально изменившиеся с тех пор.
Из лохковского века Шотландии – 410–414 млн л. н. – известны наземные многоножки Albadesmus almondi, Pneumodesmus newmani, Cowiedesmus eroticopodus, Archidesmus macnicoli и Palaeodesmus tuberculata. Они обладали первыми дыхальцами и трахейной системой дыхания. Изобилие видов явно свидетельствует об успехе группы. Вместе с тем становится ясно, чем питались первые наземные скорпионы и зачем они вообще полезли из воды на сушу.
Маленькая тонкость
Шотландские отложения с многоножками изначально ошибочно были датированы границей венлокской и лудловской эпох, то есть нижнего и позднего силура, но в последующем датировка была исправлена, так что выход многоножек на сушу несколько отложился, хотя всё равно остался в силуре.
Раннедевонская шотландская Rhyniognatha hirsti долгое время считалась древнейшим летающим насекомым, но на самом деле тоже оказалась многоножкой.
Наконец, в девоне возникают шестиногие Hexapoda. Это ещё не были настоящие насекомые, но уже очень похожие на них создания. На роль предков насекомых претендовали многие существа. Один из лучших кандидатов – раннедевонский морской Wingertshellicus backesi (он же Devonohexapodus bocksbergensis) с датировкой 405 млн л. н. – вытянутая фасеточно-пучеглазая зверюшка с усиками, у которой были шесть длинных ног спереди, ногоподобные выросты на каждом членике тела и две церки на конце тела. Впрочем, происхождение насекомых из морских или уже наземных существ ещё предстоит доказать.
Гораздо ближе к насекомым, а раньше даже включались в них ногохвостки Collembola. Древнейшим представителем была Rhyniella praecursor с древностью 410 млн л. н.
Как вы там, потомки?
Современные ногохвостки – чрезвычайно примитивные шестиногие, часто выделяемые в отдельный класс. Древность и неприхотливость позволяет ногохвосткам жить везде, где есть хотя бы какая-то почва с лишайниками и мхами, даже в Арктике и Антарктиде, а многим хватает просто поверхности воды. Особое внимание привлекает полное отсутствие даже зачатков крыльев и строение глаз (у кого они есть) – это скопления простых глазков, но не достигшие уровня фасеточных. Несмотря на примитивность и приближенность к предкам насекомых, ногохвосткам тоже есть чем удивить: на заднем конце тела у некоторых их них есть «прыгательная вилка», отталкиваясь которой они могут прыгать не только по земле, но даже по воде. Большинство ногохвосток выглядят довольно уныло, но некоторые весьма красивы – целиком в ярко-красных пупырышках, очень похожие на ягоды малины. Показательно, что подавляющая часть ногохвосток питается только споровыми растениями, уже существовавшими в девоне, лишь сминтуры Sminthuridae, похожие на помесь клеща и блохи, распробовали высшие растения.
Ногохвостки малы, зато чрезвычайно многочисленны, так что играют важнейшую роль в почвообразовании, в том числе утилизации трупов крупных живых существ, даже людей. Вероятно, они были одними из первых сухопутных падальщиков на планете.
Великое событие середины девона – появление насекомых Insecta. Первые насекомые известны по крайне эфемерным остаткам: буквально пара безымянных ошмётков из эмсского века Канады принадлежала первичнобескрылым щетинохвосткам Archaeognatha (они же Microcoryphia, или Machilida), а кусочки кутикулы из живетского века Гилбоа могли быть частями чешуйниц Zygentoma (или Lepismatida). Как бы невзрачны ни были эти обрывки, они отмечают новую веху в эволюции членистоногих. Как обычно, великие предки великой группы выглядели как блёклые козявочки без внятных опознавательных знаков. Триумф насекомых был ещё впереди.
Маленькая тонкость
Один из Великих Законов эволюции – закон невзрачности Великих Предков. Всё самое крутое появляется из самого убогого. Эволюция самых значимых групп живых организмов – это эволюция серости, а самые эффектные живые существа радуют глаз очень недолго, не получая развития.
Принцип прост: если у существа есть какие-то экзотические специализации, то это, конечно, прикольно и весело, но ставит крест на дальнейшем многообразии. Например, если у птиц или муравьедов исчезли зубы, то их потомкам не обрести много вариантов зубных систем, если у птерозавров и летучих мышей руки и пальцы превратились в крылья, то им трудно развить много вариантов рук, если у амфисбен и змей исчезли ноги, то им не стать танцорами. Шею жирафу, хобот слону и клыки саблезубому тигру эволюционно укоротить до обычных масштабов будет тоже сложно.
Конечно, всегда есть обходные эволюционные пути: в конце концов, когда-то у всех предков не было зубов и конечностей, но появились же, отчего бы им не возникнуть и ещё разок, да и уменьшение органов происходило неоднократно. Однако практика показывает, что существа с зашкаливающим значением каких-то особенностей обычно быстро вымирают при изменении условий.
С другой стороны, любая новая крутость начинается с какой-то чудной специализации. Например, вышедшие на сушу рыбы были далеко не самыми банальными рыбами, их лёгкие были очень странной чертой; перья, одевшие птиц, и уж тем более шерсть, покрывшая зверообразных рептилий, были крайне нетипичны для рептилий. Любой ароморфоз – переход на новый уровень эволюции, позволяющий освоить новый класс экологических ниш и выражающийся в перестройке многих систем организма – возможен только в очень нестандартных условиях, как правило – при значительной смене среды обитания и потрясениях экосистемы.
Всё великое возникает из простого, но начинается с хитрого приподвыверта. Диалектика – она и в эволюции диалектика.
* * *Всё это изобилие членистоногой нечисти стало возможным благодаря развитию наземных растений. Для раннего девона у нас не так много свидетельств их разнообразия, но мы знаем, что тогда уже существовали грибы хитридиомицеты Chytridiomycota – паразиты растений. Стало быть, было на ком паразитировать. Всё из того же шотландского местонахождения Райни, столь богатого на примитивнейшие сосудистые растения и многоножек, происходят раннедевонские грибы Palaeomyces, Palaeoblastocladia и Glomites.
Особое внимание привлекает растение Aglaophyton major из пражского века Шотландии. Во-первых, его строение почти идеально промежуточно между мхами и сосудистыми растениями (несмотря на то, что сосудистые явно возникли раньше; видимо, аглаофитон уже в девоне был «живым ископаемым»): с одной стороны, у него не было корней, как у мхов, с другой – спорофит в жизненном цикле преобладал, как у сосудистых растений. Во-вторых, аглаофитон обладал древнейшей арбускулярной микоризой, то есть симбиотическими грибами в межклетниках, от которых он получал всё хорошее, в частности – вечно дефицитный фосфор.
Как вы там, потомки?
Симбиоз грибов с растениями бывает разным. Регулярно он скатывается в паразитизм, причём, что удивительно, победителем тут может быть и растение. Особого стиля жизни растения – микогетеротрофы, когда растения получают все питательные вещества от грибов, вообще переставая фотосинтезировать. Примерами могут служить бесхлорофильные цветочки Thismia rodwayi и Voyria tenella, питающиеся за счёт грибов Glomeromycota. Дальше всех по этой скользкой дорожке зашла орхидея Gastrodia pubilabiata из Японии и с Тайваня: мало того, что она паразитирует на мицелии грибов, так ещё и опыляется дрозофилами, летящими на запах грибов, чтобы отложить в них яйца. Гастродия пахнет как грибы, и её белёсые цветы выглядят как шляпки поганок, глупые мухи обманываются и опыляют растение, а личинки, вылупившиеся из отложенных яиц, гибнут от голода. Так что гастродия умудряется навредить всем, к тому же и выглядит довольно мерзко, как и полагается записному злодею.
Тонкости строения позволяют рисовать детальную эволюционную преемственность сосудистых растений. К раннедевонским Horneophytopsida относится, например, Tarrantia; эта группа, возможно, является альтернативой всем остальным сосудистым растениям и не имеет современных потомков. Крайне примитивно выглядит Rhynia gwynne-vaughanii – вероятный общий предок всех сосудистых из Шотландии, чуть продвинутее Cooksonia; линию развития продолжает китайский Eophyllophyton bellum и ещё более продвинутый Psilophyton. Далее следуют уже растения, иногда уверенно определяемые как плауновидные Lycopodiophytina (или Lycopodiophyta), или их непосредственные предки – Zosterophyllopsida, у которых спорангии располагались по бокам стеблей: Zosterophyllum, более прогрессивный Protobarinophyton obrutchevii и ещё более совершённая Nothia aphylla. Тут уже начинаются совсем настоящие плауны Lycopodiopsida (или Lycopsida), в том числе шотландский Asteroxylon mackei, и вездесущий во всём Северном полушарии Drepanophycus, иногда выделяемый в самостоятельный отряд Drepanophycales. Эти соракасантиметровые растения уже имели филлоиды – подобие листьев, в которые, правда, не заходили сосуды, а также длинные – до 20 см – ризомы, независимо возникшие аналоги корней.
Для неспециалиста, конечно, все эти и многие-многие другие раннедевонские растения выглядят «на один стебель», но растения развивались лавинообразно. Если в лохковском веке они были представлены по сути теми же прибрежными рогульками, что и в силуре, то уже с пражского века – примерно с 411 млн л. н. – становятся разнообразными, суша покрывается первыми нормальными зарослями.
Enigmophyton superbum из раннего девона Красноярска и границы среднего и позднего девона Шпицбергена был первым растением с большими веерообразными листьями, несколько похожими на листья гинкго, хотя план строения всё ещё крайне примитивен.
Плауны захватывают планету. Примитивными, но успешными были Protolepidodendrales, например Protolepidodendron scharyanum. Leclercqia uncinata из живетского века Китая была покрыта довольно заковыристыми колючками. Поскольку защищаться в это время вроде бы было ещё не от кого, колючки с крючочками, скорее всего, были нужны для цепляния за другие растения, то есть леклеркия была первой лианой. Стало быть, было за что цепляться! Позднедевонский плаун Cyclostigma kiltorkense был уже приличным деревом метров в пять высотой.
В среднем и тем более позднем девоне деревья резко увеличиваются в размерах. Широко распространяется особый класс Cladoxylopsida (или Cladoxylales), что-то среднее между хвощами и папоротниками; вплоть до середины карбона они были едва ли не главными растениями суши. Самые древние, хотя ещё и невысокие деревья на планете появились в эмсском веке: Protohyenia janovii из Сибири, Foozia minuta из Бельгии, а также множество видов Pseudosporochnus по всему северному полушарию. Однако гораздо больше разрекламированы в качестве древнейших деревьев Eospermatopteris textillis из США – 385–386 млн л. н., практически идентичные синхронным бельгийским Wattieza givetiana. В местонахождении Гилбоа около Нью-Йорка сохранились прямые стволы восьмиметровой длины, на верхушке ветвившиеся странным безлистным веником. Здорово, что этот странный лес состоял из целых трёх ярусов растений: кроме эосперматоптерисов, там росли древовидные плауны пониже, а «подлесок» формировали протосеменные растения, главная часть которых скрывалась под землёй, а на поверхность выходили фотосинтезирующие загогулины.
Другим отличным примером кладоксилопсид (или кладоксилеевых) является Duisbergia mirabilis – неветвящийся столб, верх которого был покрыт густой щёткой почти листьев. Подробно изучены стволы Xinicaulis lignescens из франского века Китая, детали строения которых сохранились благодаря быстрой консервации вулканическим пеплом. Ткани таких деревьев оказались устроены весьма хитроумно: сосуды водопроводящего слоя ксилемы формировали ветвящуюся сетку по поверхности, оставляя середину полой; низ ствола под тяжестью верха проседал и расширялся в виде луковицы, достигая диаметра до 70 см; кроме того, по поверхности ствола от почвы до верхушки извивались тонкие корни. Такие башни могли вырастать до 8 – 12 м. Самый типичный род – Cladoxylon – кустарник, существовавший с середины девона до начала карбона. В кладоксилеевых включают и группу Hyeniales (или Hyeniopsida; например, Hyenia), раньше рассматривавшихся как примитивнейшие хвощи, а также Iridopteridales с единственным средне-позднедевонским родом Ibyka, возможным предком хвощей.
Совсем настоящие папоротники появились в живетском веке от Тувы до Шпицбергена в лице Protocephalopteris praecox, они составляют группу Zygopteridales (или Coenopteridiales).
Настоящие хвощи известны с начала позднего девона, это клинолистниковые Sphenophyllales, например, Asterocalamitopsis, Pseudoborniales – Pseudobornia ursina, а также Asterocalamitales – Asterocalamites.
С середины девона распространяются и самые продвинутые протосеменные растения Progymnospermopsida (они же Progymnospermophyta, или Protopteridiales), отличными примерами которых являются Aneurophyton germanicum и Protopteridium rossicum. Древнейшие остатки этих деревьев найдены в живетском ярусе Алтая и Ненецкого автономного округа, соответственно, а в последующем – до конца девона – они росли по всему Северному полушарию. Отлично сохранились спорангии Runcaria heinzelinii, росшей 385–387 млн л. н. на территории Бельгии: в центре ажурной конструкции из завитых спиралью волокон лежит макроспора с толстой оболочкой, а по бокам её прикрывают широкие листья. Это уже очень похоже на устройство настоящего семени. Иногда протосеменных относят напрямую к папоротникообразным, особое сходство с которыми им придавали спирально закрученные молодые листья-вайи. Более продвинутые позднедевонские Archaeopteridales (или Archaeopteridophyta), например, Archaeopteris и Callixylon, очень похожи на древовидные папоротники высотой до 10 м; иногда они тоже включаются в протосеменных.
Дальнейшее развитие растений мы видим на примере Moresnetia zalesskyi – самого древнего настоящего семенного растения из позднего фаменского века Бельгии и Хакасии, и синхронной Archaeosperma arnoldii из Пенсильвании, а также других подобных растений – Glamorgania gayerii, Elkinsia polymorpha, Latisemenia longshania, Kerryia mattenii и Xenotheca devonica. С них можно начинать отсчитывать голосеменные растения. На концах тонких многократно двоящихся веточек сидели «колоски» или «бокалы» с длинными остями, внутри которых хранились огромные мегаспоры-семязачатки, становившиеся после оплодотворения полноценными семенами. Такие вместилища семян мало похожи на шишки, но по сути это именно они. Характерно, что в тех же отложениях в Хакасии найдены и древнейшие настоящие семена, названные Samaropsis.

