
Полная версия:
Палеонтология антрополога. Три эры под одной обложкой
Кистепёрые Crossopterygii сами по себе в простейшей версии подразделяются на вымерших пресноводных рипидистий Rhipidistia (они же Rhipidistiiformes, или Osteolepides) и морских целакантов Coelacanthi, представленных единственным современным отрядом Coelacanthiformes (они же Coelacanthida, или Actinistia). Как обычно, в современных классификациях всё хитрее; иногда кроссоптеригии и рипидистии воспринимаются как синоним, а целаканты оказываются независимой от них группой в ранге от отряда до класса. Рипидистии возникли раньше и имели более примитивные черты, но именно это позволило им эволюционировать в наземных четвероногих.
Двоякодышащие Dipnoi (или Dipneustomorpha; при включении сюда же Porolepiformes вместе они называются Dipnomorpha) в древности были крайне разнообразны, первые из них ещё сохраняли подвижность черепной коробки, но довольно быстро утеряли эту особенность; также у современных двоякодышащих исчез верхний дентиновый (косминовый) слой чешуи. Современные двоякодышащие представлены двумя отрядами – рогозубовых Ceratodontiformes и чешуйчатников Lepidosirenoformes.
Разные группы челюстноротых появились почти синхронно в раннем силуре.
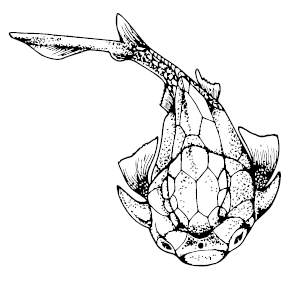
Qilinyu rostrata
Плакодерм в силуре известно не так уж много, причём большей частью по фрагментам. Из лудловской эпохи Чехии до нас дошли фрагменты артродир Kosoraspis peckai и Radotina. Этим же временем датируются целые панцири Qilinyu rostrate и чуть более продвинутого двадцатисантиметрового Entelognathus primordialis из Юннани. Эти находки особенно важны: c артродирами их роднят форма мозговой коробки и тела в целом, вытянутый туловищный панцирь и отсутствие зубов во рту, с хрящевыми рыбами и акантодами – сочленение челюстей, а с костными рыбами – строение верхней челюсти. Таким образом, эти рыбки занимают почётное место очередного Великого Предка, будучи предшественницами всех основных групп продвинутых челюстноротых; правда, величие несколько страдает от более раннего обнаружения более древних и продвинутых рыб из США и Чехии, но от тех известны лишь чешуйки, а тут – рыба в полный рост, так что китайские палеонтологи, конечно, гордо проигнорировали варварские находки и заявили о величии китайского предка.
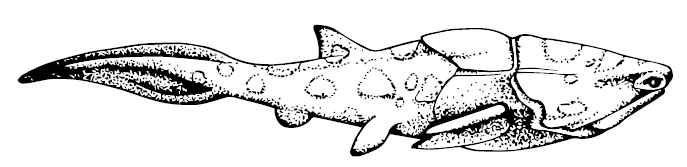
Entelognathus primordialis
Хрящевые рыбы силура известны в основном по изолированным зубам, так что трудно сказать, на что они были похожи. Древнейшие известные хрящевые рыбы – Elegestolepis grossi и E. conica, они жили от аэронского века до лудловской эпохи в морях Тувы, Алтая и Монголии. В раннесилурийском море Монголии плавали Mongolepis rozmanae, шейнвудским веком датируются тоже монгольские Deltalepis magna, D. parva и канадский Frigorilepis caldwelli, гомерским – канадский Kannathalepis. Описаны и другие роды и виды, так что ясно – в середине силура акулы уже были разнообразны. Здорово, что даже самые ранние, хотя и выделяются в свой отряд Elegestolepidida, до крайности напоминают современных акул.
Акантоды появляются в конце лландоверийской эпохи США и в венлокской Чехии. Древнейшие чешуи и плавниковые шипы не дают представления об облике их владельцев, так же как и чешуя Nostolepis striata из пржидольской эпохи Эстонии, Швеции и Германии; уже в начале девона, в лохковском веке, его сменил Nostolepis gracilis. Были, конечно, и другие силурийские акантоды – Climatius, Onchus, Gomphodus и прочие, частично переходящие в девон, где от них есть уже полноценные отпечатки.
Самые древние костные рыбы с признаками панцирных известны по фрагментам челюстей с зубоподобными пупырышками из лудловской эпохи Швеции – Andreolepis hedei и пржидольской Германии – Lophosteus superbus. Иногда этих рыб относят к палеонискам, но такое определение довольно спорно. Фрагменты настолько малы, что их можно изучать исключительно под микроскопом, а лучше – с помощью микротомографии; забавно, что для пущей контрастности на древние челюсти напыляют золото – палеонтология может быть довольно затратным занятием.
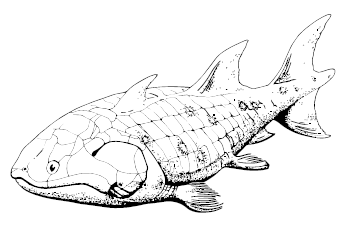
Guiyu oneiros
Позднелудловской эпохой, то есть лудфордским веком, датируется целая компания Великих Предков из Юннани, похожие находки есть и во Вьетнаме.
Из них наиболее древние и примитивные Guiyu oneiros и Sparalepis tingi из конца лудловской эпохи: в этих тридцатисантиметровых рыбках сочетались признаки хрящевых, акантод, лопастепёрых и лучепёрых: их тела были покрыты прямоугольной ганоидной чешуёй, в основании двух спинных плавников лежали здоровенные шипы, имелись зачатки окостеневших поясов конечностей, но в строении черепа признаки лопастеперых всё же перевешивали.

Sparalepis tingi
Синхронны, но более прогрессивны челюсти Megamastax amblyodus. Их уже уверенно можно отнести к лопастепёрым. Самое интересное, что мегамастакс достигал 1 м длины, будучи, таким образом, самой большой рыбой своего времени. Судя по сочетанию мелких острых и округлых пупырчатых зубов, это был хищник, способный съесть и кого-то с твёрдым панцирем. Ясное дело, это послужило поводом отрапортовать об обнаружении древнейшего хищного позвоночного, но, как часто бывает, это работает, если игнорировать более древних американских и чешских акул.
Эволюционную эстафету перенял Psarolepis romeri, живший от конца лудловской эпохи до конца лохковского века девона. Он тоже сочетал с лопастепёростью признаки лучепёрых, а потому его можно считать ещё одним почти идеальным основателем всех костных рыб. У этой хищной рыбы на конце нижней челюсти торчали крупные клыки, чем он похож на девонскую рипидистию Onychodus, перед двумя спинными и грудными плавниками располагались большие шипы, как у акантод и некоторых поздних акул (хотя как раз у примитивных акул таких шипов не было).
* * *Завоевание суши продолжалось.
Конечно, в таком ответственном деле не обошлось без странностей. Самыми загадочными образованиями, существовавшими с конца силура до конца девона, были прототакситы Prototaxites loganii. Палеонтологически они предстают как огромные окаменевшие столбы до 8,8 м высотой и 1,37 м в диаметре – это самые большие организмы своего времени! Прототакситы имеют слоистую структуру, хотя не очень понятно, была ли она строго концентрической или спиральной. На срезах видно, что ткань «ствола» состояла из каких-то волокон со сгущениями, но однозначно сопоставить такое строение с какими-то конкретными современными организмами не получается. Было выдвинуто множество предположений о природе прототакситов: они реконструировались как деревья типа плаунов, гигантские водоросли, грибы или лишайники. Не исключено, что над безжизненными равнинами силурийских побережий там и сям торчали огромные гладкие столбы. Правда, не очень понятно, как бы водоросли или грибы таких титанических размеров держались в вертикальном положении безо всякой механической ткани.
Согласно самому красивому предположению, прототакситы вообще не были единым организмом. Дело в том, что суша была заселена в лучшем случае лишайниками и мхами – организмами без корней, не способными закрепить почву. Сообщества мхов-печёночников, цианобактерий, грибов и водорослей росли в виде тонкого ковра прямо поверх песка около неустойчивых берегов. После ливней верхний край такого ковра мог загнуться, после чего под собственной тяжестью, смачиваемый водой, заворачивался в рулон. Такой скатанный мат плюхался в воду, погружался на дно, поднявшаяся муть оседала поверх, консервируя от всяких поедателей – и вот готов прототаксит. Потому-то прототакситы и исчезают в девоне – ведь именно в это время появляются нормальные наземные растения с корнями, которые хорошо держатся сами, закрепляют почву, создают постоянные берега водоёмов и своим частоколом не дают скручиваться всяким моховым коврам. Главное же подтверждение этой версии – сугубо гистологическое: микрочастицы в составе прототакситов больше всего напоминают клетки печёночников, а также там явно есть и грибные гифы, и включения цианобактерий.
Важнее, что в конце силура появились зелёные харовые водоросли Charophyta, а от их наземных микроскопических представителей или каких-то зелёных водорослей Chlorophyta возникли первые сосудистые растения Tracheophyta. По другим версиям, предками сосудистых могли стать печёночники Marchantiophyta или антоцеротовые мхи Anthocerotophyta.
Как вы там, потомки?
Харовые водоросли – удивительные существа. Внешне они очень похожи на хвощи – тот же членистый стебель, из узлов которого расходятся мутовки тонких не то веточек, не то листьев, а к земле они крепятся бесцветными вроде бы корешками. Но нет! «Междоузлия» харовых – это огромные клетки, иногда длиной в пять сантиметров, а «узлы» – стопки мелких клеточек; «корни» – на самом деле ризоиды, не имеющие свойств настоящих корней и ничем, кроме цвета, не отличающиеся от остальных частей таллома. Да к тому же взрослое растение – гаплоидный гаметофит, а не диплоидный спорофит, как у сосудистых растений. Диплоидной во всём жизненном цикле харовых является только одна клетка – зигота.
Несмотря на специфику общего строения и органов размножения, харовые водоросли – одни из лучших кандидатов на роль предков сосудистых растений. В частности, у них есть фикопласт – перегородка, возникающая при делении клетки пополам, подозрительно похожая на фрагмопласт с таким же назначением у сосудистых растений. Когда клетка делится, самый неприятный момент – «чпок» – когда мембрана двух новых клеток уже разорвалась, но у каждой дочерней ещё не схлопнулась. В этот ответственный миг из клетки может выпасть что-нибудь очень ценное. Фикопласт и фрагмопласт – как раз крышечки, появляющиеся заблаговременно и аккуратно перегораживающие разделяющиеся клетки, не дающие им развалиться как попало. Именно регуляция роста клеточной стенки – сначала для более успешного деления – стала залогом появления сосудистых растений, ведь сосуды как раз и отличаются мощной клеточной стенкой. Генетика указывает, что среди харовых самыми близкими родичами сосудистых могут быть конъюгаты, или сцеплянки Conjugatophyceae (или Zygnematophyceae), хотя современные представители этого класса очень сильно упрощены.
Маленькая тонкость
Проводящая система сосудистых растений подобна фонтану: по совокупности сосудов – ксилеме-древесине в центре растения – вода с минеральными веществами из почвы поднимается вверх к листьям, а по совокупности ситовидных элементов – флоэме-вторичной коре растения – вода с растворёнными сахарами спускается вниз от листьев к корням.
Главное достижение сосудистых, очевидно, – сосуды. Растения научились убивать некоторые свои клетки и делать из них отличные водопроводные трубы, в которых транспорту воды с минералами не мешают цитоплазма, органоиды и ядра. Сосуды позволяют закачивать воду от земли к верхушке растения. Наверху растения имеются устьица – отверстия, через которые вода испаряется, за счёт чего создаётся тяга.
Между прочим, до сих пор физика закачивания воды до конца неясна, ведь вода не просто поднимается вверх, а ещё и используется для жизни клеток; получается, испаряют они не так много, а засасывают куда как больше. Как стометровая секвойя поднимает массу воды на безумную высоту, испаряя чуток через хвою – загадка. Откуда в некоторых тропических лианах берётся такое давление, что, переруби их мачете, вода хлещет, как из шланга? Если бы ботаников так же финансировали, как физиков-ядерщиков, то у нас давно был бы бесплатный водопровод…
Сосуды бывают двух основных типов. Трахеиды имеют отверстия по бокам, а их размеры сравнительно невелики – 0,1 – 10 мм. Отдельные трахеиды появились ещё у мхов, но совершенства достигли именно у сосудистых растений. Трахеи не имеют боковых отверстий, а открыты только на концах; они достигают даже 10 см длины. Настоящие трахеи возникли только у гнётовых и покрытосеменных, причём самые продвинутые покрытосеменные, например орхидеи, остались только с трахеями, лишившись трахеид (правда, из-за этого у орхидей затруднён горизонтальный транспорт, а потому они никогда не бывают толстыми и уж подавно – деревьями).
Если же сосуды имеют к тому же толстые стенки, то получается отличная механическая ткань – каркас, дающий прочность и не позволяющий телу согнуться и упасть. Общий итог – растение с сосудами и механической тканью может стать высоким и торчать вертикально, возвышаясь над соседями и получая больше света для фотосинтеза. С этого момента начинается безумная гонка растений за место под Солнцем, не прекращающаяся и поныне.
Ситовидные элементы флоэмы, в отличие от сосудов – полуживые клетки. Дело в том, что сахара, качаемые от листьев вниз, могут заполимеризоваться, их надо растворять обратно, а для этого нужны ферменты, какая-никакая биохимия. У плаунов, хвощей, папоротников и голосеменных ситовидные клетки снабжены ядром, заострены и имеют отверстия – ситовидные поля – по бокам. У покрытосеменных ситовидные трубки уже без ядра и с ситовидными полями на концах. Для их функционирования к ним прилагаются клетки-спутницы с ядром и множеством митохондрий, обеспечивающих энергию для транспорта.
Второе принципиально важное свойство сосудистых – доминирование спорофита в жизненном цикле. Взрослое сосудистое растение диплоидно. Гаплоидная стадия гаметофита ужимается у плаунов, хвощей и папоротников до недолгого и несложного заростка с гаметангиями, а у семенных растений ещё больше – до единичных клеток, почти как у животных, так что разница между спорой и гаметой практически исчезает.
Кто бы ни был предком сосудистых, к началу силура они, видимо, уже существовали, что мы знаем по находкам характерных спор с трёхлучевой щелью разверзания в плотной оболочке из лландоверийской эпохи и даже, как уже говорилось, из позднего ордовика. Позже споры к тому же становятся разнообразны. Оледенения границы ордовика и силура могли способствовать выживанию самых упорных и подтолкнуть эволюцию растений.
Самые ранние сосудистые растения, найденные в виде отпечатков стеблей, – позднесилурийские риниофиты Rhyniophyta (они же Rhyniopsida, псилофиты Psilophyta, Psilophytites, или прапапоротникообразные Propteridophyta). Выглядели они чрезвычайно просто: тонкие веточки, регулярно разделяющиеся надвое, с пумпочками-спорангиями на концах; иногда одна ось оказывалась потолще, а ответвление – потоньше, изредка встречалось деление натрое. Самое главное – у риниофитов были настоящие сосуды, устьица, оболочка кутикула, защищающая от высыхания, и корнеподобные ризоиды в почве. В большинстве случаев риниофиты имели высоту 20–70 см, очень редко вымахивали аж до 3 м, но такие гиганты существовали уже в девоне. Риниофиты росли вдоль побережий, частично в воде, но некоторые – и вполне на суше, причём образовывали довольно плотные заросли.
Работа над ошибками
Часто термины «псилофиты» и «риниофиты» используют как синонимы, но, если начать занудствовать, между ними есть разница. Термин «Psilophytites» изначально был придуман в 1952 году для обозначения непонятных стеблей без спорангиев («псилос» – голый), не поддающихся точной классификации, то есть заранее понимался как сборная солянка. Термин же «Rhyniophyta» с самого начала означал определённую систематическую группу примитивнейших сосудистых растений. Назван род Rhynia был в честь шотландской деревни Райни, около которой были обнаружены первые отпечатки; однако в русской литературе прижилась транскрипция «риния», а для всей группы, соответственно, «риниофиты».
Замечательно, что ещё до фактического открытия окаменелостей (что случилось в 1912 г.) риниофиты были успешно предсказаны в 1908 г. французским ботаником О. Линье, который, исходя из чистой логики, достаточно детально описал Propsiloteae, гипотетических предков сосудистых растений. И ведь О. Линье угадал все ключевые свойства. Вот она – предсказательная сила науки!
Самая древняя и примитивная представительница риниофитов – Cooksonia – известна по отпечаткам стеблей как минимум с венлокской эпохи; впрочем, некоторые считают её слишком примитивной даже для риниофитов. Эти маленькие растения были устроены проще некуда: несколько раз ветвящиеся вилочки не выше пяти сантиметров со спорангиями на верхушках. Самый древний вид – Cooksonia barrandei – найден в Чехии и датирован 432 млн л. н.; такие же датировки у неназванных куксоний из Ирландии. Показательно, что, с одной стороны, один и тот же род обнаруживается в синхронных отложениях по всему миру – Европе, Северной Америке, Западной Сибири, а с другой стороны, уже в конце силура известно довольно много видов, отличающихся деталями строения. Таким образом, куксония – отличный великий предок всех сосудистых растений планеты.
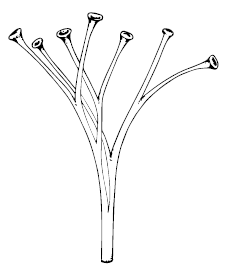
Cooksonia
В конце силура по берегам водоёмов уже густо колосились многочисленные сосудистые растения – Yarravia oblonga, Ya. subsphaerica, Hedeja corymbosa и прочие. Некоторые из них сохраняли ещё очень примитивные черты, например, Horneophyton, Caia, Salopella и Tortilicaulis имели чрезвычайно простые сосуды и даже иногда выделяются в самостоятельный класс Horneophytopsida. Возможно, гаметофит этих растений всё ещё был тоже крупным, выглядящим примерно как и спорофит. Если это действительно так, то хорнеофитопсиды абсолютно уникальны, занимая строго промежуточное место между низшими и высшими растениями.
Наряду с очень примитивными понемногу разгоняются и более продвинутые: Baragwanathia уже отрастила многочисленные почти листья микрофиллы, в которые даже заходили сосуды, а потому больше походила на плаун, в класс каковых часто и включается. Такие растения известны от горстийского века силура до эмсского века девона и от Австралии и Китая до Чехии и Канады.
Как вы там, потомки?
В современной флоре существуют странные растения – псилотовидные Psilotales (или Psilotopsida) – Psilotum nudum и Tmesipteris elongata. Внешне они чрезвычайно похожи на силурийские и раннедевонские растения. У псилота точно такое же дихотомическое деление стебля, нет ни листьев, ни корней, на верхушках находятся спорангии; собственно, псилофиты и получили свое название за сходство с псилотом. Долгое время ботаники спорили – примитивны ли псилотовидные исходно. Это дожившие до нас или же упростившиеся вторично псилофиты. То есть их предки были в некоторый момент уже совсем даже ничего, но потом стали снова похожи на куксонию, в буквальном смысле слова скуксились. Решение пришло из генетики: псилотовидные оказались ужовниковыми папоротниками Ophioglossidae, опсилофитившимися до силурийского состояния. Не берите с них пример – кистепёрыми станете!
Если на берегах появляется много растительных остатков, то кому-то это понравится. Видимо, уже в конце силура на суше распространились грибы, питающиеся древесиной. Жизнь подготавливалась к великому завоеванию суши, которое в полной мере свершилось уже в следующем – девонском – периоде.
* * *Как уже говорилось, вторая половина силура выдалась жаркая и засушливая. В потеплевших водах океана кислород растворялся не очень хорошо, и растения, и животные океана чувствовали себя как в супе и потихоньку мёрли. Конец силура ознаменовался сокращением разнообразия морских организмов, в том числе многих брахиопод. Впрочем, это вымирание не идёт ни в какое сравнение с предыдущим, так что в девон перешли почти все группы.
* * *Силур – поворотный период нашей истории. Он дал нам челюсти и конечности, а самое главное – уверенность в грядущем. Под гнётом ракоскорпионов выковывалось недалёкое доминирование позвоночных. Одновременно, не теряя времени, первые сосудистые растения начали уверенное освоение суши – потомки куксоний сыграли принципиальную роль в становлении разума, дав нашим предкам и кров, и стол, и топливо для очага.
АльтернативыСилур – последний период, когда членистоногие имели шанс развить разум и захватить планету. Ракоскорпионы были крупнейшими тварями своего времени; жизнь в пограничных условиях – на грани солёной и пресной воды, воды и суши – могла бы простимулировать их развитие. Но, как многократно случалось в эволюции, первопроходцы попались в ловушку чересчур экстремальных условий и одновременно отсутствия конкуренции. Возникшие скорпионы оказались настолько хорошо приспособлены к жизни на пустынной суше, что навеки остановились в развитии и до сих пор выглядят неотличимо от своих силурийских предков.
Девон
419,2–358,9 миллиона лет назад: Лето палеозоя, время рыб и Великий Десант
МЕЖДУНАРОДНАЯ И РОССИЙСКАЯ ШКАЛА:
419,2 млн л.н. – ранний девон: лохковский век – 410,8 – пражский век – 407,6 – эмсский век – 393,3 – средний девон: эйфельский век – 387,7 – живетский век – 382,7 – поздний девон: франский век – 372,2 – фаменский век – 358,9

Девон – один из самых важных периодов для нашего становления. Длиной почти со всю кайнозойскую эру, он просто не мог не быть богат на события. Период, когда суша оказалась окончательно и бесповоротно завоевана всеми теми, кто живёт на ней поныне, – по масштабу преобразований это время сопоставимо с кембрием. Девон принял Землю с камнями, а оставил с лесами.
* * *Огромная Гондвана из слившихся Южной Америки, Африки, Антарктиды и Австралии на Южном полюсе, Северная Америка, Северная Европа, Казахстан и Китай на уровне экватора, выстроившиеся дугой, окружающей океан Палеотетис, Сибирь, как полагается Сибири – на севере, океан Панталасса на половину планеты – карта девона радует взор.
Климат девона – один из теплейших за всё время существования планеты, по температурам на третьем месте после кембрия и эоцена. В первую половину периода стояла жара и сушь, в том числе по всей территории Гондваны, во второй половине стало холодать, так что в конце фаменского века случилось даже оледенение.
Морская жизнь в девоне испытывала некоторый застой: в первую половину периода разнообразие морских животных ещё росло, а потом потихоньку стало снижаться. Впрочем, бедным девон назвать уж точно невозможно.
В девоне впервые появились известковые губки Calcarea (возможно, они были уже в силуре, но это не точно). Продолжался расцвет кораллов табулят.
Как часто бывает, после насыщения вариантами строения началась гонка размеров. Большие существа часто имеют преимущества перед всякой мелочью. Жертвы становятся больше, быстрее и защищённее, хищники отращивают когти-челюсти-открывашки. Девон – время, когда эти тенденции проявились наилучшим образом. Отличный пример – гигантский многощетинковый червь Websteroprion armstrongi с границы раннего и среднего девона Канады. От этой полихеты сохранились лишь крючковатые асимметричные челюсти длиной 10–15 мм, но сравнение с современными аналогами показывает, что длина тела была около 1 м.
Если есть такие страшные хищники – будут и соответствующие жертвы. Среди брахиопод очень показателен Echinaurus из Марокко, чья раковина покрыта множеством длинных тонких игл.
Как вы там, потомки?
Современные хитоны – совершенно замечательные моллюски. Снизу у них одна сплошная нога, как и положено улиткам, но сверху раковина разделена на шесть последовательных сегментов, чем очень напоминает спину мокриц, причём первый и последний щитки отличаются от шести средних. По бокам тела идут пары жабр, сегментированы и панцирные мускулы; нормальной мантийной полости нет; нервная система без ганглиев, нет также статоцистов, глаз и щупалец; анальное отверстие расположено на конце тела, как у червей. Все эти особенности, конечно, всегда интерпретировались как свидетельство чрезвычайной архаичности хитонов; регулярно эти животные упоминаются как «переходное звено» от червей к моллюскам. Конечно, нынешние хитоны – никакое не «промежуточное звено», но вся группа явно очень рано отделилась от прочих моллюсков и, возможно, действительно сохранила некоторые черты общих с червями кембрийских предков. Тем более удивительно, что первые достоверные хитоны известны лишь из девона.
Среди моллюсков появляются хитоны, или панцирные моллюски Polyplacophora (они же Polyplaxiphora, или Loricata); североамериканский Strobilepis spinigera является древнейшим достоверным представителем класса, хотя не раз в качестве хитонов описывались и кембрийские, и ордовикские раковинки.

