 Полная версия
Полная версияEssays Upon Heredity and Kindred Biological Problems
Now however minute the units may be, there is not the least doubt that the limits of possible division have been long since reached by all existing species, for we may safely assume that no one of them has acquired the sexual method of reproduction within a small number of recent generations. All existing species must therefore now contain as many different kinds of ancestral germ-plasms as they are capable of containing; and the question arises,—How can sexual reproduction now proceed without a doubling of the quantity of germ-plasm in each germ-cell, with every new generation?
There is only one possible answer to such a question:—sexual reproduction can proceed by a reduction in the number of ancestral germ-plasms, a reduction which is repeated in every generation.
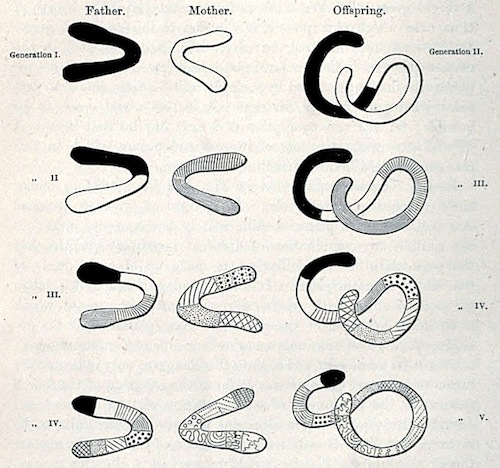
Fig. I.
This must be so: the only question is, how and when does the supposed reduction take place.
Inasmuch as the germ-plasm is seated, according to our theory, in the nucleus, the necessary reduction can only be produced by nuclear division; and quite apart from any observation which has been already made, we may safely assert that there must be a form of nuclear division in which the ancestral germ-plasms contained in the nucleus are distributed to the daughter-nuclei in such a way that each of them receives only half the number contained in the original nucleus. After Roux’s256 elaborate review of the whole subject, we need no longer doubt that the complex method of nuclear division, hitherto known as karyokinesis, must be considered not merely as a means for the division of the total quantity of nuclear substance, but also for producing a division of the quantity and quality of each of its single elements. In by far the greater number of instances the object of this division is obviously to effect an equal distribution of nuclear substance in the two daughter-nuclei, so that each of the different qualities contained in the mother-nucleus is transferred to the two daughter-nuclei. This interpretation of ordinary karyokinesis is less uncertain than perhaps at first sight it may appear to be. We cannot, it is true, directly see the ancestral germ-plasms, nor do we even know the parts of the nucleus which are to be looked upon as constituting ancestral germ-plasm; but if Flemming’s original discovery of the longitudinal division of the loops lying in the equatorial plane of the nuclear spindle is to have any meaning at all, its object must be to divide and distribute the different kinds of the minutest elements of the nuclear thread as equally as possible. It has been ascertained that the two halves produced by the longitudinal splitting of each loop never pass into the same daughter-nucleus, but always in opposite directions. The essential point cannot therefore be the division of the nucleus into absolutely equal quantities, but it must be the distribution of the different qualities of the nuclear thread, without exception, in both daughter-nuclei. But these different qualities are what I have called the ancestral germ-plasms, i.e. the germ-plasms of the different ancestors, which must be contained in vast numbers, but in very minute quantities, in the nuclear thread. The supposition of a vast number is not only required by the phenomena of heredity but also results from the comparatively great length of the nuclear thread: furthermore it implies that each of them is present in very small quantity. The vast number together with the minute quantity of the ancestral germ-plasms permit us to conclude that they are, upon the whole, arranged in a linear manner in the thin thread-like loops: in fact the longitudinal splitting of these loops appears to me to be almost a proof of the existence of such an arrangement, for without this supposition the process would cease to have any meaning.
This is the only kind of karyokinesis which has been observed until recently; but if the supposed nuclear division leading to a reduction in the number of ancestral germ-plasms has any real existence, there must be yet another kind of karyokinesis, in which the primary equatorial loops are not split longitudinally, but are separated without division into two groups, each of which forms one of the two daughter-nuclei. In such a case the required reduction in the number of ancestral germ-plasms would take place, for each daughter-nucleus would receive only half the number which was contained in the mother-nucleus.
Now there is more evidence for the existence of this second kind of karyokinesis than the fact that it is demanded by my theory; for I believe that it has been already observed, although it has not been interpreted in this sense.
It is very probable that this is true of van Beneden’s257 observation on the egg of Ascaris megalocephala: he found that the nuclear division which led to the formation of the polar body differs from the ordinary course of karyokinesis, in that the plane of division is at right angles to that usually assumed. Carnoy258 has confirmed this observation in its main features, and he has made the further observation that out of the eight nuclear loops which are found at the equator of the spindle, four are removed with the first polar body, and that half of the remaining four are removed with the second polar body. The first of these two divisions would have to be looked upon as a reduction, if it is certain that each of the eight nuclear loops consists of different ancestral germ-plasms; but this assumption is impossible, although on the other hand it cannot be directly disproved: for we are not able to see the ancestral germ-plasms. But it must nevertheless be maintained that the removal of the first four loops does not imply a reduction in the number of ancestral germ-plasms in the nucleus; because, as I have already argued, two successive divisions of the number of ancestral germ-plasms into halves is inconceivable; and because the first polar body is also present in parthenogenetic eggs in which such division into halves cannot take place. But the karyokinetic process can readily be looked upon as a removal of ovogenetic nucleoplasm, for we know from the observations of Flemming and Carnoy, that, under certain circumstances, subsequent divisions may occur, involving an increase in the number of nuclear loops to double their number. These subsequent divisions of course take place in the daughter-nuclei. This fact proves, as I think, that there are nuclei in which the same ancestral germ-plasm occurs in two different loops: but such loops, identical as regards the composition of their ancestral germ-plasms, may very well contain different ontogenetic stages of this substance. This will be the case in the instance alluded to, if four loops of the first nuclear spindle are to be looked upon as ovogenetic nucleoplasm, and the four others as germ-plasm. It is therefore unnecessary to regard the first division of the egg-nucleus as a ‘reducing division’: it may be looked upon as an ‘equal division’259 entirely analogous to the kind of division which, in my opinion, directs the development of the embryo. This conclusion would receive direct proof if it were possible to show that the eight loops of the first division have arisen by the longitudinal splitting of four primary loops: for a longitudinal splitting of the nuclear thread would be the means by which the different ontogenetic stages of the germ-plasm could be separated from one another, without leading to any reduction in the number of ancestral germ-plasms in the daughter-nuclei. Thus I have previously attempted to prove that the ontogenetic development of the egg must be connected with a progressive transformation of the nucleoplasm during successive nuclear divisions, and this transformation will very frequently (but not always) occur in such a way that the different qualities of the nucleoplasm are separated from one another by the nuclear division. The nucleoplasm of the daughter-nuclei will be identical if the two daughter-cells are to potentially contain corresponding parts of the embryo; as for instance the first two segmentation spheres of the egg of the frog, which according to Roux260 correspond to the right and left halves of the future animal. But the nucleoplasm must be unequal if the products of division are to develope into different parts of the embryo. In both cases, however, karyokinesis is connected with a longitudinal splitting of the nuclear threads, and we may conclude from this fact (which is also confirmed by the phenomena of heredity) that all such nuclei, whether they have entered upon the same or different ontogenetic transformations of their nucleoplasm, are identical as regards the ancestral germ-plasm which they contain. During the whole process of segmentation and the entire development of the embryo, the total number of ancestral germ-plasms which were at first contained in the germ-plasm of the fertilized egg-cell must still be contained in each of the succeeding cells.
Thus no objection can be raised against the view that the four loops of the first polar body contain the ovogenetic nucleoplasm, that is to say, an idioplasm which contains the total number of ancestral germ-plasms, but at an advanced and highly specialized ontogenetic stage.
The formation of the second polar body may be rightly considered as a ‘reducing division,’ as a division leading to the expulsion of half the number of the different ancestral germ-plasms, in the form of two nuclear loops, for no reason can be alleged in support of the assumption that the four loops of the second nuclear spindle are made up of identical pairs. Furthermore the facts of heredity require the assumption that the greatest possible number of ancestral germ-plasms is accumulated in the germ-plasm of each germ-cell, and thus that the small number of loops not only means an increase in quantity but a multiplication in the number of different ancestral germ-plasms present in each of them. If this conclusion be correct, there can be no doubt that the second division of the egg-nucleus means a reduction in the above-mentioned sense.
But there are yet other observations which, if correct, must also be considered as ‘reducing divisions.’ I refer to all those cases in which the longitudinal splitting of the loops is either entirely wanting, or does not occur until after the loops have left the equator of the spindle and have moved towards the poles. In both instances the bearing upon the question would be the same, for only half the number of primary loops would reach each pole in either case. If therefore the primary loops are not made up of identical pairs, it follows that the two daughter-nuclei can only contain half the number of ancestral germ-plasms which were contained in the mother-nucleus. Whether the loops divide on their way to the poles or at the poles themselves, no difference will be brought about in the number of ancestral germ-plasms which they contain, for this number can neither increase nor diminish. The quantity of the different ancestral germ-plasms can alone be increased in this way. I am here referring to observations made by Carnoy261 on the cells which form the spermatozoa in various Arthropods. It must be admitted, however, that these divisions cannot be regarded as ‘reducing divisions,’ if Flemming’s262 suggestion be confirmed, that in all these observations the fact has been overlooked that the equatorial loops are not primary but secondary, and that they have arisen from the longitudinal splitting of the nuclear thread during previous stages of nuclear division. But this point can only be decided by renewed investigation. Although many excellent results have been obtained in the subject of karyokinesis, there is still very much to be learnt before our knowledge is complete; and this is not to be wondered at when we remember the great difficulties in the way of observation which are chiefly raised by the minute size of the objects to be investigated. Flemming’s most recent publications prove that we are still in the midst of investigation, and that highly interesting and important processes have hitherto escaped attention. A secure basis of facts is only very gradually obtained, and there are still many conflicting opinions upon the details of this process. I should therefore consider it to be entirely useless, from my point of view, to enter into a critical examination of everything known about all the details of karyokinesis. I am quite content to have shown how it may be imagined that the reduction required by my theory takes place during nuclear division; and at the same time to have pointed out that there are already observations which may be interpreted in this sense. But even if I am mistaken in this interpretation, the theoretical necessity for a reduction in the number of ancestral germ-plasms, a reduction repeated in every generation, seems to me to be so securely founded that the processes by which it is effected must take place, even if they are not supplied by the facts already ascertained. There must be two kinds of karyokinesis according to the different physiological effect of the process. First, a karyokinesis by means of which all the ancestral germ-plasms are equally distributed in each of the two daughter-nuclei after having been divided into halves: secondly, a karyokinesis by means of which each daughter-nucleus receives only half the number of ancestral germ-plasms possessed by the mother-nucleus. The former may be called ‘equal division,’ the latter ‘reducing division.’ Of course these two processes, which differ so greatly in their effects, must also be characterized by morphological differences, but we cannot assume that the latter are necessarily visible. Just as, during the division of the first and second nuclear spindle in the egg of Ascaris megalocephala, karyokinesis takes, upon the whole, the same morphological course, although we must ascribe different physiological meanings to the two processes of division,—so it may be in other cases. The ‘reducing division’ must be always accompanied by a reduction of the loops to half their original number, or by a transverse division of the loops (if such division ever occurs); although reduction can only occur when the loops are not made up of identical pairs. And it will not always be easy to decide whether this is the case. On the other hand, the form of karyokinesis in which a longitudinal splitting of the loops takes place before they separate to form the daughter-nuclei must always, as far as I can see, be considered as an ‘equal division.’ In the accompanying figures II and III, diagrams are given illustrating these two forms of karyokinesis, but I do not mean to imply that it is impossible to imagine any other form in which they may occur.
In Figure II a nuclear spindle is seen at A, and at its equatorial zone there are twelve primary loops. The transverse cross-lines and other markings on the loops indicate that they are composed of different ancestral germ-plasms. The loops are shaded differently in order to render the diagram clear. At B six of the loops are seen to have moved to either pole, so that the figure is a representation of the ‘reducing division.’ Figure III is a diagrammatic representation of ‘equal division.’ The six loops at the equatorial zone of A are shown by different cross-lining and shading to be composed of different ancestral germ-plasms. The loops split longitudinally in a direction indicated by the longitudinal line upon each of them. In B the halves of the loops are seen to have moved to the opposite poles of the spindle, so that there are not only six loops at each pole, but also all the six combinations of ancestral germ-plasms.
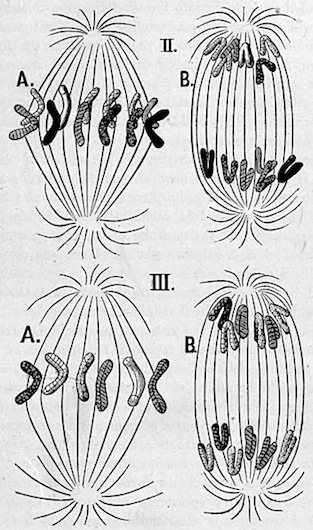
Figs. II, III.
Perhaps some may be inclined to look upon direct nuclear division as a ‘reducing division,’ but I believe that such a view would be incorrect. It is only approximately true that the nuclear thread is divided into two halves of equal quantity by direct division, and exact equality would only happen as it were accidentally; so that we cannot speak of a perfectly equal distribution of the ancestral germ-plasm in the two daughter-nuclei. But the ‘reducing division’ must obviously effect an exactly regular and uniform distribution of the ancestral germ-plasms, although this does not imply that every ancestral germ-plasm of the mother-nucleus would be represented in each of the two daughter-nuclei. But if out of e.g. eight nuclear loops at the equatorial plane, four pass into one, and the other four into the other daughter-nucleus, each of the latter will contain an equal number of ancestral germ-plasms, although different ones. This is indeed part of the foundation of the theory, for the ‘reducing division’ must remove exactly half of the original number of ancestral germ-plasms, and precisely the same number must be replaced at a later period by the sperm-nucleus. This could hardly be achieved with sufficient precision by direct nuclear division.
I now come to inquire whether the expulsion of the second polar body is in reality, as I have already maintained, a reduction in the number of ancestral germ-plasms present in the nucleus of the egg. The view itself is sufficiently obvious, and it would supply an explanation of the meaning of the process which is still greatly wanted; but it will nevertheless be not entirely useless to consider other possible theories.
It would be quite conceivable to suppose that the youngest egg-cells, which multiply by division, may undergo one ‘reducing division’ in addition to the ordinary process. Of course this should occur once only, for if repeated, the number of ancestral idioplasms in the nucleus of the germ-cell would undergo a decrease greater than could be afterwards compensated by the increase due to fertilization. Thus the number of ancestral germ-plasms would continually decrease in the course of generations,—a process which would necessarily end with their complete reduction to a single kind, viz. to the paternal or the maternal germ-plasm. But the occurrence of such a result is disproved by the facts of heredity. Although such an early occurrence of the ‘reducing division’ would offer advantages in that nothing would be lost, for both daughter-nuclei would become eggs, instead of one of them being lost as a polar body, nevertheless I do not believe that it really occurs: weighty reasons can be alleged against it.
Above all, the facts of parthenogenesis are against it. If the number of ancestral germ-plasms received from the parents were reduced to half in the ovary of the young animal, how then could parthenogenetic development ever take place? It is true that we cannot at once assert the impossibility of an early ‘reducing division’ on this account, for as I have shown above, the power to develope parthenogenetically depends upon the quantity of germ-plasm contained in the mature egg; the necessary amount might be produced by growth, quite independently of the number of different kinds of ancestral germ-plasms which form its constituents. The size of a heap of grains may depend upon the number of grains, and not upon the number of different kinds of grains. But in another respect such a supposition would lead to an unthinkable conclusion. In the first place, the number of ancestral germ-plasms in the germ-cells would be diminished by one half in each new generation arising by the parthenogenetic method; thus after ten generations only 1/1024 of the original number of ancestral germ-plasms would be present.
Now, it might be supposed that the ‘reducing division’ of the young egg-cells was lost at the time when the parthenogenetic mode of reproduction was assumed by a species; but this suggestion cannot hold, because there are certain species in which the same eggs can develope either sexually or parthenogenetically (e.g. the bee). It seems to me that such cases distinctly point to the fact that the reduction in the number of ancestral germ-plasms must take place immediately before the commencement of embryonic development, or, in other words, at the time of maturation of the egg. It is only decided at this time whether the egg of the bee is to develope into an embryo by the parthenogenetic or the sexual method; such decision being brought about, as was shown above, by the fact that only one polar body is expelled in the first case, while two are expelled in the second. But if we are obliged to assume that reproduction by means of fertilization, necessarily implies a reduction to one half of the number of ancestral germ-plasms inherited from the parents,—the further conclusion is obvious, that the second division of the egg-nucleus and the expulsion of the second polar body represent such a reduction, and that this second division of the egg-nucleus is unequal in the sense mentioned above, viz. one half of the ancestral germ-plasms remains in the egg-nucleus, the original number being subsequently restored by conjugation with a sperm-nucleus; while the other half is expelled in the polar body and perishes.
I may add that observations, so far as they have extended to such minute processes, do indeed prove that the number of loops is reduced to one half. It has been already mentioned that, according to Carnoy, such reduction occurs in Ascaris megalocephala, but the same author also describes the process of the formation of polar bodies in a large number of other Nematodes263, and his descriptions show that the process occurs in such a way that the number of ancestral germ-plasms must be reduced by half. Sometimes half the number of primary loops pass into the nucleus of the polar body, while the other half remains in the egg. In other cases, as in Ophiostomum mucronatum, the primary nuclear rods divide transversely,—a process which must produce the same effect. It is true that these observations require confirmation, and since, with unfavourable objects, the difficulties of observation are extremely great, there may have been errors of detail; but I do not think that there is any reason for doubting the accuracy of the essential point. And this essential point is the fact that the number of primary loops is divided into half by the formation of the polar body.
But even if we could not admit that such a conclusion is securely founded, it cannot be doubted that the formation of the second polar body reduces to one half the quantity of the nucleus which would have become the segmentation-nucleus in the parthenogenetic development of the egg. This is a simple logical conclusion from the two following facts: first, parthenogenetic eggs expel only one polar body; secondly, there are eggs (such as those of the bee) in which it is absolutely certain that the same half of the nucleus—which is expelled as the second polar body in the egg requiring fertilization—remains in the egg when it is to develope parthenogenetically, and acts as half of the segmentation-nucleus. But this proves that the expelled half of the nucleus must consist of true germ-plasm, and thus a secure foundation is laid for the assumption that the formation of the nucleus of the second polar body must be considered as a ‘reducing division.’
I was long ago convinced that sexual reproduction must be connected with a reduction in the number of ancestral germ-plasms to one half, and that such reduction was repeated in each generation. When, in 1885, I brought forward my theory of the continuity of the germ-plasm, I had long before that time considered whether the formation and expulsion of polar bodies must not be interpreted in this sense. But the two divisions of the egg-nucleus caused me to hesitate. The two divisions did not seem to admit of such an interpretation, for by it the quantity of the nucleus is not divided into halves, but into quarters. But a division of the number of ancestral germ-plasms into quarters would have caused, as was shown above, a continuous decrease, leading to their complete disappearance; and such a conclusion is contradicted by the facts of heredity. For this reason I was led at that time to oppose Strasburger’s view that the expulsion of the polar bodies means a reduction of the quantity of nuclear substance by only half. My objection to such a view was valid when I said that the quantity of idioplasm contained in the egg-nucleus is not, as a matter of fact, reduced to one half, but to one quarter, inasmuch as two successive divisions take place. I may add that I had also considered whether the two successive divisions might not possess an entirely different meaning,—whether one of them led to the removal of ovogenetic nucleoplasm, while the other resulted in a reduction in the number of ancestral germ-plasms. But at that time there were no ascertained facts which supported the supposition of such a difference, and I did not wish to bring forward the idea, even as a suggestion, when there was no secure foundation for it. The morphological aspects of the formation of the first and second polar bodies are so extremely similar that such a supposition might have been considered as a mere effort of the imagination.

