 Полная версия
Полная версияEssays Upon Heredity and Kindred Biological Problems
These views contain many fallacies quite apart from the soundness or unsoundness of their foundation. The process of encystment, as Götte thinks, represents, in the Monoplastides, true reproduction to which multiplication by means of division has been secondarily added. This encystment cannot be dispensed with, for internal causes determine that it must occasionally interrupt the process of multiplication by simple division. But, on the other hand, Götte also considers the division of the contents of the cyst to be a secondary process. The essential characteristic of encystment is a simple process of rejuvenescence without multiplication. Hence we are forced to accept a primitive condition in which simple division as well as the division of the encysted individual were absent, and in which reproduction consisted only in an often-repeated process of rejuvenescence among existing individuals, without any increase in their number. Such a condition is inconceivable because it would involve a rapid disappearance of the species, and the whole consideration clearly shows us that division of un-encysted individuals must have existed from the first, and that this, and not a vague and mysterious rejuvenescence, has always been the real and primitive reproduction of the Monoplastides. The fact that encystment does not always lead to the division of the contents of the cyst proves, in my opinion, that not reproduction but preservation against injury from without, was the primitive meaning of encystment. It is possible that at the present time there are but few Monoplastides which are able to go through an infinite number of divisions without the interposition of the resting condition implied by encystment; although it has not yet been demonstrated for all species67. But it is not right to conclude from this that there is an internal necessity which leads to encystment, that is to say to identify this process with rejuvenescence. It is much more probable that encystment is merely an adaptation to continual changes in the external conditions of life, such as drought and frost, and perhaps also the want of food which arises from the over-population of small areas. The same phenomenon is known in certain low Crustacea—the Daphnidae—which possess an ephippium or protective case for their winter-eggs. This case is only developed after a certain definite number of generations has been run through, an event which may happen at any time in the year in species living in pools which are liable to be often dried-up; but only in the autumn in such as live in lakes which are never dry. No one ever doubted that the periodical formation of the ephippium in certain generations was an adaptation to changes in the external conditions of life.
Even if the process of rejuvenescence in the Monoplastides were really equivalent to the death of the higher animals, we could not conclude from this that it is necessarily associated with reproduction. Encystment alone is not reproduction, and it first becomes a form of reproduction when it is associated with the division of the encysted animal. Simple division was the true and original form of reproduction in Monoplastides, and even now it is the principal and fundamental form.
Hence we see that among the Monoplastides reproduction is not connected with death, even if we accept Götte’s view and allow that encystment represents death. I shall return later on to the relation between death and reproduction in the Metazoa; but the question first arises whether encystment, if it is not death, has any analogue in the higher animals, and further whether death takes that place in their development which is occupied by encystment in the Monoplastides.
Among the higher Metazoa there can be no doubt as to what we mean by death, but the precise nature of that which dies is not equally evident, and the popular conception is not sufficient for us. It is necessary to distinguish between the mortal and the immortal part of the individual—the body in its narrower sense (soma) and the germ-cells. Death only affects the former; the germ-cells are potentially immortal, in so far as they are able, under favourable circumstances, to develope into a new individual, or, in other words, to surround themselves with a new body (soma)68.
But how is it with the lowest Polyplastides in which there is no antithesis between the somatic and germ-cells, and among which each of the component cells of the multicellular body has retained all the animal functions of the Monoplastides, even including reproduction?
Götte believes that the natural death of these organisms (which he rightly calls Homoplastides) consists in ‘the dissolution of the cell-colony.’ As an example of such dissolution Götte takes Häckel’s Magosphaera planula, a marine free-swimming organism in the form of a sphere composed of a single layer of ciliated cells, imbedded in a jelly. (For figure see below.) This organism cannot however be ‘considered as a genuine perfect Polyplastid, for at a certain time the component cells part from one another and then continue to live independently in the condition of Monoplastides.’ These free amoebiform organisms increase considerably in size, encyst, and finally undergo numerous divisions—a kind of segmentation within the cyst. The result of the division is a sphere of ciliated cells similar to that with which the cycle began. In fact, Magosphaera is not a perfect Polyplastid, but a transitional form between Polyplastides and Monoplastides, as the discoverer of the group of animals of which it is the only representative, indicated, when he named the group ‘Catallacta.’
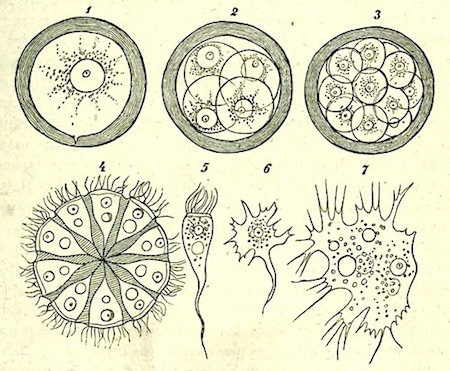
Development of Magosphaera Planula (after Häckel).
1. Encysted amoeboid form. 2 and 3. Two stages in the division of the same. 4. Free ciliated sphere, the cells of which are connected by a gelatinous mass. 5. One of the ciliated cells which has become free by the breaking up of the sphere. 6. The same in the amoeboid form. 7. The same grown to a larger size.
According to Götte, the natural death of Magosphaera consists, as in the undoubted Protozoa, in a process of rejuvenescence by encystment. The dissolution of the ciliated sphere into single cells ‘cannot be identical with natural death. For the regular and complete separation of the Magosphaera-cells proves that their individuality has not been completely subordinated to that of the whole colony, and it proves that the latter is not completely individualised69.’
Nothing can be said against this, if we agree in identifying death with the encystment of the Monoplastides. Now we could, as Götte rightly remarks, derive the lower forms of Polyplastides from Magosphaera if ‘the connection between the cells of the ciliated sphere were retained until encystment, viz. until the reproduction of the single cells had taken place70.’ After this had been accomplished, Götte considers that death would consist ‘in the complete separation of the cells from one another, accompanied in all probability by their simultaneous change into germ-cells.’ The fallacy in this is evident; if death is represented in one case by the encystment during which single cells change into germ-cells, then this must apply to the other case also, for nothing has changed except the duration of the cell-colony. The nature of encystment cannot be affected by the fact that the cells separate from one another a little earlier or a little later. If it is true that death is represented by encystment among the Monoplastides, then the same conclusion must also hold for the Polyplastides; or rather death must be represented in them by the process of rejuvenescence, which Götte considers to be the essential part of encystment. Götte ought not to identify death with the dissolution of the cell-colony of which the lowest and highest Polyplastides are alike composed; but he should seek it in the process of rejuvenescence which takes place within the germ-cells. If it is essential to the nature of reproduction that the cells set apart for that purpose should pass through a process of rejuvenescence, which is equivalent to death, then this must be true for the reproductive cells of all organisms. If these conclusions hold good, there is nothing to prevent us from assuming that such a process of rejuvenescence actually occurs in the higher animals. Götte evidently holds this view, as is plainly shown in the last pages of his essay. He there attempts to bring his views of the death and rejuvenescence of the germ into harmony with his previously developed idea of the derivation of death among the Polyplastides from the dissolution of the cell-colonies. Götte still clings to the view which he propounded in describing the development of Bombinator, according to which the egg-cell of the higher Metazoa must pass through a process of rejuvenescence representing death, before it can become a germ.
According to Götte’s71 idea ‘the egg of a Bombinator igneus before fertilization cannot be considered to be a cell either wholly or in part; and this is equally true of it at its origin and after its complete development; it is only an essentially homogeneous organic mass enclosed by a membrane which has been deposited externally.’ This mass is ‘unorganised and not living72,’ and ‘during the first phenomena of its development all vital powers must be excluded.’ In this way the continuity of life between two successive individuals is always interrupted; or, as Götte says in his last essay:—‘The continuity of life between individuals of which one is derived from the other by means of reproduction, exists neither in the rejuvenescence of the Monoplastides nor in the condition of the germ among the Polyplastides—a condition which is derived from the former73.’
This is quite logical, although in my opinion it is both unproved and incorrect. But, on the other hand, it is certainly illogical for Götte to derive the death of the Metazoa in a totally different way, i. e. from the dissolution of their cell-colonies. It is quite plain that the death of the Metazoa does not especially concern the reproductive cells, but the individual which bears them; Götte must therefore seek for some other origin of death—an origin which will enable it to reach the body (soma)—as opposed to the germ-cells. If there still remained any doubt about the failure to establish a correspondence between death and the encystment of the Monoplastides, we have here, at any rate, a final demonstration of the failure!
But there is yet another great fallacy concealed in this derivation of the death of the Polyplastides.
Among the lowest Polyplastides, where all the cells still remain similar, and where each cell is also a reproductive cell, the dissolution of the cell-colony is, according to Götte, to be regarded as death, inasmuch as ‘the integrity of the mother-individual absolutely comes to an end’ (l. c., p. 78). The dissolution of a cell-colony into its component living elements can only be called death in the most figurative sense, and can have nothing to do with the real death of the individuals; it only consists in a change from a higher to a lower stage of individuality. Could we not kill a Magosphaera by boiling or by some other artificial means, and would not the state which followed be death? Even if we define death as an arrest of life, the dissolution of Magosphaera into many single cells which still live, is not death, for life does not cease in the organic matter of which the sphere was composed, but expresses itself in another form. It is mere sophistry to say that life ceases because the cells are no longer combined into a colony. Life does not in truth cease for a moment. Nothing concrete dies in the dissolution of Magosphaera; there is no death of a cell-colony, but only of a conception. The Homoplastides, that is cell-colonies built up of equal cells, have not yet gained any natural death, because each of their cells is, at the same time, a somatic as well as a reproductive cell: and they cannot be subject to natural death, or the species would become extinct.
It is more to the purpose that Götte has sought for an illustration of death among those remarkable parasites, the Orthonectides, because in them we do at any rate meet with real death. They are indeed very low organisms; but nevertheless they stand far above Magosphaera, even if the latter were hypothetically perfected up to the level of a true Homoplastid, for the cells which compose the body of the Orthonectides are not all similar, but are so far differentiated that they are even arranged in the primitive germ-layers, and a form results which has rightly been compared with that of the Gastrula. It is true they are not quite so simple as Götte74 figures them, for they not only consist of ectoderm and germ-cells, but, according to Julin75, the endoderm is arranged in two layers—the germ-cells and a layer which forms during development a strong muscular coat; and in the second female form the egg-cells are surrounded by a tolerably thick granular tissue.

Orthonectides (after Julin).
8. First female form: the cap-like anterior part has become detached and the egg-cells (eiz) are escaping.
9. Second female form: eiz = egg-cells; outside these are the muscular layer (m) and the ectoderm (ekt).
10 and 11. Two fragments of such a female broken to pieces by spontaneous division: the egg-cells are embedded in a granular mass, and undergo embryonic development in it at a later period; the whole is surrounded by ciliated cells.
12. Male discharging the spermatozoa by the breaking up of the ectoderm (ekt); sp spermatozoa; m muscle.
There is nevertheless no doubt that in the first female form, when sexually mature, the greater part, not only of the endoderm but of the whole body, is made up of ova, so that the animal resembles a thin-walled sac full of eggs. The ova escape by the bursting of the thin ectoderm, and when they have all escaped, the thin disintegrated membrane, composed of ciliated cells, is no longer in a condition to live, and dies at once. This is the course of events as described by Götte, and he is probably correct in his interpretation. This is the real death of the Orthonectides, and if we regard them as low primitive forms (Mesozoa), here for the first time in the ascending series we meet with natural death. But the causes of this are scarcely so clear as Götte seems to think when he ascribes it to the effect of reproduction—an effect which is ‘not only empirically necessary, but absolutely unavoidable.’ Such a necessity is explained by the fact that the endoderm consists entirely of germ-cells. Now the life of the organism, being dependent upon the mutual action of both layers, must cease as soon as the whole endoderm is extruded during reproduction.
Arguments such as these pass over the presence of a mesoderm; but apart from this omission, it does not appear to me so self-evident from a purely physiological standpoint, that the ectodermal sheath with its muscle layer must die after the extrusion of the germ-cells.
In those females to which Götte refers in this passage, the whole sheath remains at first quite uninjured, with the exception of a small cap at the anterior end, which is pushed off to give exit to the ova; and inasmuch as the sheath continues to swim about in the nutritive fluids after this has taken place, the proof is at any rate wanting that it cannot support itself quite as well as before, although it has lost the germ-cells.
Then why does it die? My answer to this is simple:—because it has lived its time; because its length of life is limited to a period which corresponds with the time necessary for complete reproduction. The physical constitution of the body is so regulated that it remains capable of living until the extrusion of the reproductive cells, and then dies, however favourable external conditions may be for its further support.
The correctness of this explanation is shown by a consideration of the males and the second form of females; for in these cases the body falls to pieces, not as a consequence of reproduction, but as a preparation for it!
Götte only mentions the second female form in a note, in which he says, it appears ‘that in the second female form of these animals the whole body breaks into many pieces, and the superficial layer gradually atrophies, so that it dies before the eggs are extruded.’ In Julin’s account76, upon which Götte bases his statements, there are, however, some not unimportant differences. For instance, the eggs are not extruded at all, but embryonic development takes place within the body of the mother, which has previously undergone spontaneous division into several pieces. In this case, the eggs differ from those of the other female form, inasmuch as they do not constitute the whole of the endoderm, but are embedded (as was stated above) in a fairly voluminous granular mass at the expense of which, or at least by means of which, they are nourished; for they increase considerably in size during their development. But not only this granular mass, but all the layers of the body of the mother, even the ectoderm, persist during the embryonic development of the offspring. Indeed, the ectoderm must continue to grow during the division of the mother animal, for it gradually covers in the products of division on all sides, and, by means of its cilia, causes the animal to swim about in the fluids of its host. After some time the cilia are lost, and the separate parts into which the mother animal has divided, fix themselves upon some part of the body-cavity of the host; the young become free, and the remains of the body of the mother probably disappear by dissolution and resorption77. In this case the remains of the mother animal seem to be, to some extent, consumed by the embryos,—a process which sometimes, although very rarely, happens elsewhere. We can scarcely consider this as a primitive arrangement, or look upon it as a proof that ‘reproduction’ has a necessarily fatal effect upon the Polyplastid organism.
In the male, the mass of spermatozoa does not swell out the body to such an extent that its walls must give way and thus permit an exit, but the large ectoderm cells atrophy spontaneously at the time of maturity, and as they fall off, exit is given to the spermatozoa here and there. In this instance also the dissolution of the body is not a consequence of reproduction, but reproduction can only take place when the dissolution of the body has preceded it!
In this remarkable arrangement we cannot discern anything except an evident adaptation of the life of the body-cells to reproductive purposes, and this adaptation was rendered possible because, after the evacuation of the sexual cells, the body ceased to be of any value for the maintenance of the species.
But even if we assume, that the death of the Orthonectides is, in Götte’s sense, a consequence of reproduction, inasmuch as, in the two forms of females as well as in the male, the extrusion of a mass of developed germ-cells or embryos deprives the organism of the physiological possibility of living longer, how can we explain the necessity of death as a direct consequence of reproduction in all Polyplastides? Is the body—the soma—of the Metazoa so imperfectly developed, as compared with the reproductive cells, that the extrusion of the latter involves the death of the former? As a matter of fact in the majority of cases the relations are reversed; the number of body-cells usually exceeds the germ-cells a hundred- or a thousand-fold, and the body is, as regards nutrition, so completely independent of the reproductive cells, that it need not be in the least disadvantageously affected by their extrusion. And if the Orthonectid-like ancestors of the Metazoa were compelled to give up their insignificant somatic part after the extrusion of their germ-cells, because it could now no longer support itself, does it therefore follow that the somatic cells had for ever lost the power of surviving, even when their phyletic descendants were surrounded by more favourable conditions? Had they to inherit ‘the necessity of death’ for all time? Whence came this great change in the nature of organisms which, before the differentiation of Homoplastids into Heteroplastids, were endowed with the immortality of unicellular beings?
And it must be remembered that it is only an assumption which places the Orthonectides among the lowest Metazoa (Heteroplastids). I do not intend to greatly emphasize this point, but the formation of the Gastrula by embole, and the absence of a mouth and alimentary canal, shows that these parasites are extremely degenerate, and the same may be said of almost all endoparasites. The Gastrula, as an independent organism, was without doubt primitively provided with both mouth and stomach, and the mass of ova filling the female Orthonectid is an adaptation to a parasitic life, which on the one side renders the possession of a stomach a superfluity, and on the other demands the production of a great number of germ-cells78. It is certain that the Orthonectides, as at present constituted, cannot have lived in the free condition, and also that their adaptation to parasitism cannot have arisen at the beginning of the phyletic development of Metazoa, because they inhabit star-fishes and Nemertines—both relatively highly developed Metazoa. Hence it is, at any rate, doubtful whether the Orthonectides can claim to pass as typical forms of the lowest Heteroplastids, and whether their reproduction can be looked upon ‘as typical for the unknown ancestors of all Polyplastids’ (l. c., p. 45). If, however, we accept some organism resembling these Orthonectides as the most ancient Heteroplastid, being a free-living organism, it must have had a stomach, and the cells surrounding it must—as a whole or in part—have possessed the power of digesting; at any rate, they cannot all have been germ-cells, and therefore it is improbable that death would be the direct result of the extrusion of the germ-cells.
Let us now consider the manner in which Götte has endeavoured to explain the transmission of the cause of death—which first appeared in the Orthonectides—from these organisms to all later Metazoa, until the very highest forms are reached. Exact proofs of this supposition are unfortunately wanting, and the evidence is confined to the collection of a number of cases in which death and reproduction take place nearly or quite simultaneously. These would prove nothing, even if post hoc were always propter hoc; and there are, opposed to them, a number of cases in which reproduction and death take place at different times. In obtaining evidence for ‘the fatal influence of reproduction,’ is it possible to point to every case of sudden death after the act of oviposition or fertilization? These cases occur among many of the higher animals, especially in Insects, and were collected by me in an earlier work79. It is obvious that such cases are exceptional, but in a restricted sense it is quite true, as far as these individual instances are concerned, that death appears as a consequence of reproduction. The male bee, which invariably dies while pairing, is undoubtedly killed in consequence of a very powerful nervous shock; and the female Psychid, which has laid all her eggs at once, dies of ‘exhaustion’—however we may attempt to explain the term on physiological principles.
Can we conclude from these cases that the effects of reproduction are, in Götte’s sense, universally fatal; that reproduction is the positive and ‘exclusive explanation of natural death’? (l. c., p. 32.) I need not linger over these isolated examples, but I turn at once to the foundation of the whole conclusion—a foundation which is obviously unable to support the superstructure erected on it. Götte formally derives the idea that death is a necessary condition of reproduction, from a very heterogeneous collection of facts. When we examine this collection we find that the process which is taken to be death is not the same thing in all these instances, while the same is true of the influence of reproduction by which death is supposed to be caused. The whole conception arises out of the process of encystment, which is regarded as the building-up of reproductive material—that is, as true reproduction; and since, according to Götte’s view, the formation of germs is always intimately connected with an arrest of life, and since, by his own definition, this stand-still of life is equivalent to death, it follows that, with such a theory, reproduction, in its essential nature, must be inseparably connected with death. It is necessary at this juncture to remember what Götte means by the process of rejuvenescence, and to point out that he is dealing with something quite different from ‘the fatal influence of reproduction,’ which was just now mentioned with regard to insects. ‘Rejuvenescence,’ bound up as it is with encystment and reproduction, is, according to Götte, ‘a re-coining of the specific protoplasm, by means of which the identity of its substance is fixed by heredity,’ a ‘marvellous process in which phenomena the most important in the whole life of the animal, and in fact of all organisms—reproduction and death—have their roots’ (l. c., p. 81). Whether such re-coining really takes place or not, at any rate I claim to have shown above that it does not correspond with death in the Metazoa, and—if it is represented at all in these latter—that it ought to be looked for in the reproductive cells; and indeed, in another passage, Götte himself has placed the process in these cells.

