
Полная версия:
Экология. Повреждение и репарация ДНК: учебное пособие
1.2.2. Участие корректирующих автономных экзонуклеаз в репликации и репарации ДНК
Корректирующая экзонуклеазная активность не обязательно является частью процессивной ДНК-полимеразы. Достаточно часто в клетке для коррекции новосинтезированной ДНК привлекаются многочисленные автономные экзонуклеазы. Эти экзонуклеазы способны отщеплять неправильный нуклеотид, образуя (или не образуя) комплекс с ДНК-полимеразами.
При синтезе ведущей нити основную роль играют ДНК-полимераза III Е. coli, ошибочность которой составляет 10-6, а у эукариот ДНК-полимеразы ε и δ (ошибочность 10-5). Различные автономные экзонуклеазы способны повышать точность этих процессивных полимераз в 5-10 раз.
При синтезе отстающей нити у Е. соli в объединении фрагментов Оказаки участвует ДНК-полимераза I. Точность этой умеренно процессивной полимеразы резко увеличивается в присутствии автономных экзонуклеаз.
Большинство систем репарации ДНК нуждаются в синтезе ДНК при исправлении ошибок. У Е. соli в репарации ДНК существенное участие принимает ДНК-полимераза I, точность работы которой явно увеличивается в присутствии автономных экзонуклеаз. В клетке человека за сутки образуется в среднем 105 спонтанных повреждений ДНК в результате ее дезаминирования, депуринизации, окисления и неправильного метилирования. Большинство этих повреждений репарируются при участии ДНК-полимеразы β (ошибочность 10-3), В результате этого в транскрибируемой ДНК каждый раунд репликации появляются приблизительно 3 ошибки, подлежащие коррекции. При работе другой системы репарации непроцессивные безнуклеазные сверхошибочные (ошибочность 10-2) ДНК-полимеразы ведут синтез ДНК в обход повреждений в ДНК-матрице. После завершения синтеза на поврежденной матрице они должны быть заменены, по-видимому, с помощью репликативного фактора С, на основную элонгирующую ДНК-полимеразу δ (ошибочность 10-5). Но в силу той же инерционности биохимических реакций замена не происходит мгновенно, что требует исправления ошибок (совершенных непроцессивными полимеразами напротив неповрежденной матрицы), в том числе с участием автономных экзонуклеаз. В экстрактах клеток человека недавно показана экзонуклеазная коррекция ошибок, допущенных сверхошибочной ДНК-полимеразой η (эта). Наконец, автономные экзонуклеазы участвуют в коррекции гетеродуплексов.
Все виды экзонуклеазной коррекции должны закончиться за время данной репликации. По-видимому, коррекция ДНК-полимеразных ошибок – весьма эффективный процесс, поскольку анализ генома человека показал, что дивергенция последовательностей в транскрибируемой ДНК составляет примерно 0,1 % при исследовании ДНК от 24 человек различных этнических групп.
2. Типы повреждений ДНК
Большинство повреждений ДНК не являются результатом только ошибок репликации. Множество повреждений возникает в любое время клеточного цикла под действием как экзогенных, так и эндогенных факторов. В табл. 1. схематично изображены основные типы повреждений ДНК, которые опознаются и устраняются различными системами репарации.
Ультрафиолетовые лучи вызывают образование пиримидиновых димеров, 6,4-фотопродуктов, аддуктов, разрывов и прочие повреждения ДНК. Под действием химических агентов происходят разного рода модификвции нуклеотидов, возникают межнитевые сшивки, конформационные дефекты. Двунитевые разрывы могут приводить к перестройкам хромосом, что и является главной причиной летального действия ионизирующей радиации.
Таблица 1. Основные типы повреждений ДНК.
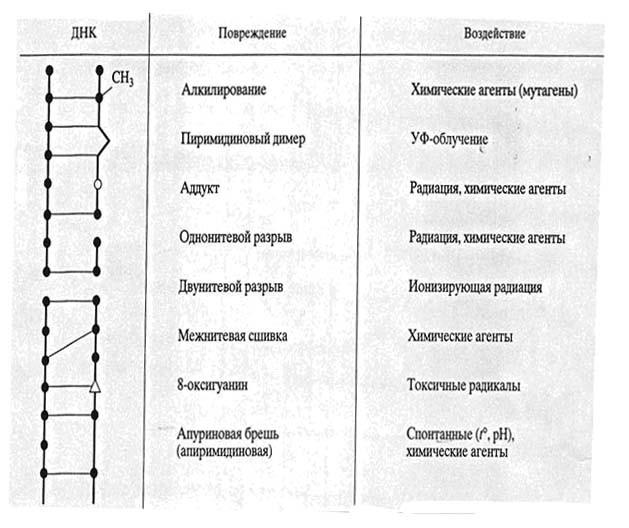
В результате внутриклеточных процессов в ДНК образуются многочисленные АP-сайты из-за спонтанной утраты пуринов/пиримидинов, окисленные участки (например, 8-оксигуанин), возникающие под действием токсических радикалов, постоянно генерируемых в процессах метаболизма.
Следует заметить, что все повреждения показаны только схематически, так как любое из них вызывает локальное изменение структуры ДНК вокруг, а разрывы ДНК почти всегда сопровождаются и модификацией прилежащих к ним оснований. Кроме обычно упоминаемых пиримидиновых димеров, под действием УФ облучения образуются и другие повреждения, например, 6,4-фотопродукты. В дальнейшем, описывая различные системы репарации ДНК, мы будем подробно останавливаться на тех типах повреждений, которые они способны исправлять.
3. Многообразие систем репарации ДНК
Системы репарации ДНК крайне разнообразны – от простых одноэтапных (фотореактивация, деалкилирование) до сложнейших, многоэтапных механизмов, контролируемых большим числом генов и, включающих соответственно, большое число белков. Этим репарация принципиально отличается от процессов репликации и рекомбинации, обходящихся существенно более ограниченным набором биохимических реакций. Множественность частично перекрывающихся и дополняющих друг друга систем репарации, даже некоторая их избыточность, повышает надежность защиты генома и расширяет возможности обеспечения его работы в онтогенезе и при различных физиологических условиях. Классификация процессов репарации ДНК строится на тех реакциях, которые являются их основой. Выделяются реакции прямой репарации, системы эксцизионной репарации ДНК, репарация с привлечением рекомбинации, пострепликативная репарация, репарация двунитевых разрывов, репаративный обход повреждения, причем иногда один и тот же тип репарации может иметь несколько разных названий. В результате исследований последних лет эта классификация менялась и уточнялась, что мы в дальнейшем будем учитывать.
Конец ознакомительного фрагмента.
Текст предоставлен ООО «ЛитРес».
Прочитайте эту книгу целиком, купив полную легальную версию на ЛитРес.
Безопасно оплатить книгу можно банковской картой Visa, MasterCard, Maestro, со счета мобильного телефона, с платежного терминала, в салоне МТС или Связной, через PayPal, WebMoney, Яндекс.Деньги, QIWI Кошелек, бонусными картами или другим удобным Вам способом.
Вы ознакомились с фрагментом книги.
Для бесплатного чтения открыта только часть текста.
Приобретайте полный текст книги у нашего партнера:
Полная версия книги
Всего 10 форматов

