Полная версия:
Физиология силы
Особый интерес представляет механизм, лежащий в основе автоматических шагательных движений. Согласно современным представлениям, в процессе шагания существенное значение принадлежит спинальному генератору шагательных движений (В.С. Гурфинкель и др., 1998; Ю.П. Герасименко, 2000). Предполагается, что интернейронный ансамбль, реализующий шагание, состоит из объединенных в кольцо обособленных функциональных групп. Возбуждение распространяется по кольцу, поочередно включая одну группу мышц за другой. Однократное распространение возбуждения по кольцу обеспечивает реализацию функционального элемента шагания – шаг одной и шаг другой ноги. Такой цикл многократно повторяется. Включение шагания и регуляция его в конкретных условиях обеспечиваются согласованной деятельностью сегментарных и надсегментарных образований.
Афферентные входы мотонейронного пула. Регуляция скелетных мышц в целом и ДЕ как их составных частей осуществляется не только со стороны надсегментарных двигательных систем и спинномозговых интернейронных образований, но и за счет афферентных сигналов, идущих к спинальным а-мотонейронам от рецепторного аппарата самих мышц, суставов и окружающей их кожной поверхности. К этим рецепторам относятся: мышечные веретена, сухожильные рецепторы, суставные рецепторы и механорецепторы кожи (рис. 1.6).
Мышечные веретена информируют о длине мышцы и скорости изменения длины при ее сокращении. Они располагаются продольно ходу мышечных экстрафузальных (вневеретенных) волокон (рис. 1.6). Мышечное веретено представляет собой сложное инкапсулированное структурное образование длиной 4-10 мм, включающее 3 структуры: 10–15 внутриверетенных (интрафузальных) волокон, рецепторные окончания и двигательный аксон.
В зависимости от расположения ядер различают два типа интрафузальных мышечных волокон: с ядрами, сгруппированными в средней части веретена в виде сумки (ЯС-волокна), и с ядрами, расположенными цепочкой вдоль веретена (ЯЦ-волокна). Интрафузальные волокна обоих типов спиралевидно оплетены окончаниями афферентных нервных волокон, по которым и передается информация в спинной мозг. Чувствительность интрафузальных волокон регулируется гамма-мотонейронами, тела которых располагаются в передних рогах спинного мозга, а их длинные отростки (аксоны) иннервируют сами интрафузальные волокна.
Волокна типа ЯС функционально делятся на динамические и статические. Первые реагируют преимущественно на скорость изменения длины мышцы, вторые – на изменение длины вне зависимости от скорости ее изменения. Интрафузальные волокна типа ЯЦ являются статическими.

Рис. 1.6. Схема афферентных аппаратов мышц (Л.О. Бадалян, И.А. Скворцов, 1986)
1 – афферент 1b; 2 – афферент 1а; 3 – афферент II; 4 – альфа-мотонейрон; 5 – гамма-мотонейрон; 6 – нервно-мышечное веретено; 7 – сухожильный рецептор; 8 – экстрафузальное мышечное волокно; 9-10 – интрафузальные мышечные волокна с ядерной сумкой (9) и ядерной цепочкой (10)
Сухожильные рецепторы сигнализируют о степени напряжения мышцы при ее сокращении. Они располагаются в области соединения сухожилия и мышцы. Один конец рецептора прикрепляется к сухожилию, а другой – вплетается в интерстиций мышцы.
Рецептор имеет веретенообразную форму, а в его подкапсулярном пространстве располагаются чувствительные нервные окончания, реагирующие на сокращение мышцы. От сухожильного рецептора к спинному мозгу направляются афферентные нервные волокна 1b. При сокращении мышцы рецептор растягивается в продольном направлении и генерирует импульсы в спинной мозг. Чем больше напряжение мышцы, тем большая импульсация направляется в спинной мозг. Импульсы, идущие по 1b афферентам, оказывают влияние на мотонейроны собственной мышцы через полисинаптические цепи с участием тормозных нервных клеток.
Суставные рецепторы информируют о величине и скорости изменений суставного угла, а также о направлении движения в данном суставе. Суставные рецепторы находятся преимущественно в суставных сумках и связках. Они разделяются на медленно и быстро адаптирующиеся. Первые сигнализируют о суставном угле, т. е. о положении сустава. При увеличении угла частота афферентных импульсов возрастает. Быстро адаптирующиеся рецепторы информируют о скорости движения в суставе. При этом, чем больше скорость движения, тем чаще импульсация в афферентных нервных волокнах.
Механорецепторы кожи модифицируют временные и амплитудные характеристики проприоцептивных двигательных рефлексов и поэтому играют заметную роль в регуляции произвольных движений (Р.М. Городничев, 1987).
Произвольный контроль движений. Регуляция движений посредством рефлексов является простейшей формой управления целенаправленными двигательными действиями. Координация произвольных спортивных движений, т. е. осуществляемых при участии сознания спортсмена, базируется на активности высших центров головного мозга: коры, мозжечка, базальных ганглиев. Эти структуры мозга инициируют движение и корректируют его в ходе непосредственного выполнения.
Кора головного мозга. Разные корковые области определяют целесообразность двигательных действий, регулируют положение тела в пространстве, создают двигательные программы и корректируют их в различных ситуационных условиях, а также обеспечивают включение элементарных движений в сложные поведенческие реакции человека. Управляющие сигналы из коры головного мозга направляются по нисходящим нервным путям к мотонейронам спинного мозга.
Основные нисходящие тракты к мотонейронам спинного мозга начинаются в коре больших полушарий и стволе мозга. В зависимости от области серого вещества спинного мозга, которого они достигают, эти тракты (пути) разделяются на две группы – латеральные и медиальные. Латеральные тракты включают латеральный кортикоспинальный тракт, начинающийся в коре мозга, и руброспинальный тракт, начинающийся в красном ядре среднего мозга. Медиальные пути включают кортикоспинальный тракт, вестибулоспинальные тракты, ретикулоспинальные тракты и тектоспинальный тракт. Тракты называются по тем участкам мозга, с которого они начинаются и на каком из них заканчиваются.
Латеральные двигательные пути. Латеральный кортикоспинальный тракт начинается в моторной и премоторной областях коры больших полушарий. Его волокна оканчиваются на интернейронах и мотонейронах латеральной части спинного мозга, контролирующих дистальные мышцы, которые обеспечивают тонкие движения пальцев. Руброспинальный тракт начинается в красном ядре и заканчивается на интернейронах и мотонейронах, связанных с латеральной двигательной системой. Нейроны красного ядра имеют возбуждающие входы от моторной зоны коры и мозжечка. Предполагается, что руброспинальный путь дублирует ряд функций кортикоспинального тракта. После повреждения обоих латеральных двигательных путей значительно нарушается координация движений верхних и нижних конечностей.
Медиальные двигательные пути. Медиальные пути начинаются преимущественно в стволе мозга и направляют свои волокна к мотонейронам медиальной части спинного мозга, которые иннервируют проксимальную группу скелетных мышц. Аксоны нейронов вестибулоспинального тракта формируют возбуждающие моносинаптические входы на мотонейронах мышц-разгибателей, и активируемые через интернейроны спинного мозга тормозные входы на мышцы-сгибатели. Функция вестибулоспинального тракта состоит в регуляции положения тела в пространстве и активации мышц-разгибателей, противодействующих влиянию гравитации (силе тяжести). Ретикулоспинальные тракты обеспечивают двустороннее возбуждение медиальных мотонейронов разгибателей и торможение мотонейронов, регулирующих проксимальные конечности. Тектоспинальный тракт отвечает за движения головы и глаз, вызываемые зрительными и слуховыми раздражителями.
Кора головного мозга в реализации произвольных движений. Координация двигательной деятельности происходит при непосредственном участии нейрональных сетей коры головного мозга. Важное значение имеет активность нейронов первичной моторной коры – М1 и вторичной моторной коры, состоящей из премоторной зоны и дополнительной моторной зоны (рис. 1.7).
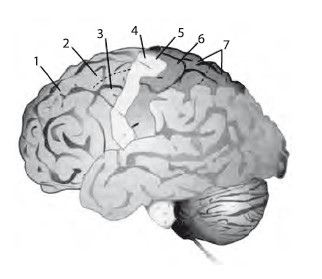
Рис. 1.7. Кора головного мозга
1 – лобная доля,
2 – дополнительная моторная кора,
3 – премоторная кора,
4 – первичная моторная кора,
5 – центральная борозда,
6 – сенсорная кора,
7 – теменная доля
Моторная кора организована соматотопически, т. е. активность определенного участка коры приводит к сокращению мышцы, располагающейся в строго определенной области тела (рис. 1.8). При этом мышцы в коре представлены не пропорционально: проекции мышц лица и рук более обширны по сравнению с мышцами туловища. Анализ мозга человека методом позитронно-эмиссионной томографии выявил, что активность нейрональных сетей моторной коры связана с выполняемыми движениями (Дж. Г. Николлс и др., 2008).

Рис 1.8. Представительство мышц в коре головного мозга
1 – глотательных, 2 – языка, 3 – челюстных, 4 – губ, 5 – лица, 6 – глазодвигательных, век, 7 – надбровья, 8 – шеи, 9 – большого пальца, 10 – указательного пальца, 11 – среднего пальца, 12 – безымянного пальца, 13 – мизинца, 14 – ладони, 15 – запястья, 16 – предплечья, 17 – плеча, 18 – туловища, 19 – бедра, 20 – голени, 20 – стоп, 21 – пальцев ног
Результаты регистрации нейрональной активности моторной коры показали строгую зависимость возможности генерации разрядов, их частотных характеристик, а также особенностей временных связей между разрядами активных нейронов от параметров выполняемого двигательного действия.
Оказалось, что часть кортикальных нейронов активна в процессе сгибания, другие же – в процессе разгибания. Одни нейроны активизировались в начале движения, а другие – при его завершении. Частота разрядов нейронов повышалась по мере возрастания мышечного усилия. Траектория движения в пространстве связана с максимальной частотой разрядов определенной группы кортикальных нейронов. Планирование и координация двигательных действий являются весьма сложными процессами, поэтому кроме коры головного мозга в регуляции движений существенное значение имеет активность и других мозговых структур, в частности – мозжечка.
Мозжечок. Функции мозжечка во многом определяются его связями с другими структурами нервной системы. Он обладает массивными проекциями от моторной коры и получает информацию от проприорецепторов, находящихся в мышцах и суставах, а также от рецепторов зрительной и вестибулярной сенсорных систем. Мозжечок участвует в регуляции равновесия и контроле положения тела. В специальных исследованиях была доказана роль мозжечка в инициации и исполнении планируемых двигательных действий.
Мозжечок контролирует рефлексы, вызываемые активацией мышечных веретен, посредством чего достигается плавность сокращения скелетных мышц. Важное значение имеет интегративная деятельность мозжечка по сравнению достигнутого двигательного результата с планируемым и внесению в случае установления дисбаланса (несоответствия) соответствующих коррекционных поправок. В качестве примера можно рассмотреть такое произвольное движение, как удар по футбольному мячу (рис. 1.9).
Управляющие нервные импульсы из высших моторных центров передаются через латеральные и медиальные пути к соответствующим мышцам и активируют их. Импульсы также одновременно посылаются в мозжечок. В процессе удара проприорецепторы (мышечные веретена, сухожильные и суставные рецепторы) посылают сигналы о параметрах сокращения рабочих мышц обратно в мозжечок. Нейрональные сети мозжечка сравнивают полученную от проприорецепторов информацию с планируемым результатом и, если достигнутый результат не соответствует планируемому, направляют коррекционные импульсы в моторную кору. В таком случае высшие моторные центры вносят поправку в выполняемое движение или корректируют программу последующего повторного выполнения удара по мячу.
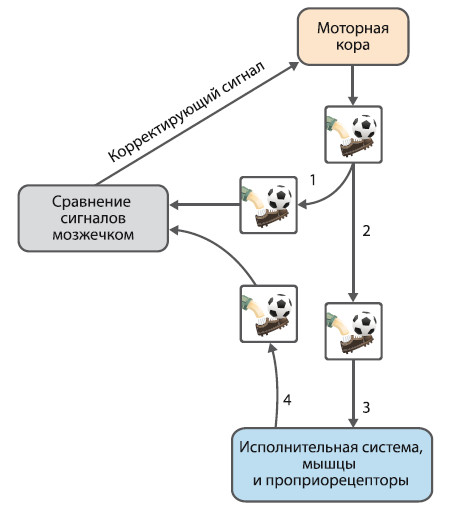
Рис. 1.9. Пример интегративной деятельности мозжечка 1 – нисходящий сигнал из моторной коры в мозжечок,
2 – нисходящий сигнал по пирамидальному тракту,
3 – нисходящий сигнал от мотонейронов к мышцам,
4 – афферентный проприорецептивный сигнал к мозжечку
Базальные ганглии. Базальные ганглии представляют собой скопление групп нейронов, объединенных в крупные ядра. Они находятся в белом веществе, рядом с дальними слоями коры больших полушарий. Базальные ганглии состоят из хвостатого ядра и скорлупового ядра, которые имеют множественные входы от нейронов, располагающихся в коре головного мозга. Нейрональная активность базальных ганглиев тормозит нейроны таламокор-тикального пути до момента поступления из моторной коры сигнала отмены. В соответствии с современными представлениями, базальные ганглии участвуют в инициации движений, поддержании позы и мышечного тонуса (К.Б. Шаповалова, 2015).
Энграммы. Многократно повторяемое движение заучивается и хранится в головном мозге в виде энграммы. В основе формирования энграмм лежат функциональные и структурные изменения нейронов и синапсов мозга, вызываемые систематически выполняемыми двигательными действиями. При необходимости энграмма реализуется в соответствующем движении. В то же время энграмма конкретного движения может быть извлечена из памяти человека и использована при осуществлении других двигательных действий.
Энграммы чрезвычайно быстрых движений хранятся в моторной зоне головного мозга и квалифицируются как двигательные программы. Они могут осуществляться без обратной сенсорной связи. Энграммы, хранимые в сенсорной области головного мозга, реализуются в медленных двигательных действиях при участии обратной афферентной информации.
Глава 2
Структура и функции нервно-мышечной системы
Знания о строении и функциях нервно-мышечной системы способствуют пониманию процессов, лежащих в основе развития силовых возможностей человека. В конечном итоге такие знания позволяют более осознанно и целеустремленно подходить к планированию и организации тренировочного процесса, нацеленного на развитие силовых способностей лиц разного пола и возраста.
Нервно-мышечный аппарат. Целенаправленная двигательная деятельность обеспечивает взаимодействие организма с внешней средой (Р.С. Персон, 1976; 1987; Ю.Т. Шапков, 1984; В.С. Гурфинкель и др., 1999; J.H. Wilmore, D.L. Kostil, 2004). Функция движения реализуется скелетными мышцами, которые сокращаются в ответ на электрические импульсы, приходящие к ним по длинным отросткам (аксонам) от а-мотонейронов – нервных клеток, располагающихся в передних рогах спинного мозга. Мышцы и иннервирующие их мотонейроны составляют нервно-мышечный аппарат человека. Сократительная деятельность мышц обеспечивает поддержание позы человека, перемещение частей тела относительно друг друга и передвижение человека в пространстве. Мышцы человека представлены в Приложении 1.
Основным морфофункциональным элементом нервно-мышечного аппарата является двигательная единица (ДЕ). ДЕ – это мотонейрон и иннервируемая им группа мышечных волокон. На рис. 2.1 схематически изображено строение ДЕ.
Аксон мотонейрона из спинного мозга проходит в составе периферического нерва до мышцы, внутри которой разветвляется на множество концевых веточек. Каждая концевая веточка заканчивается на одном мышечном волокне, образуя нервно-мышечный синапс. Эфферентные импульсы, идущие по аксону от клетки тела мотонейрона к мышце, активируют все иннервируемые им мышечные волокна. Поэтому ДЕ функционирует как единое морфофункциональное образование.

Рис. 2.1. Схематическое строение двигательной единицы (рисунок С.М. Иванова)
Конец ознакомительного фрагмента.
Текст предоставлен ООО «ЛитРес».
Прочитайте эту книгу целиком, купив полную легальную версию на ЛитРес.
Безопасно оплатить книгу можно банковской картой Visa, MasterCard, Maestro, со счета мобильного телефона, с платежного терминала, в салоне МТС или Связной, через PayPal, WebMoney, Яндекс.Деньги, QIWI Кошелек, бонусными картами или другим удобным Вам способом.
Вы ознакомились с фрагментом книги.
Для бесплатного чтения открыта только часть текста.
Приобретайте полный текст книги у нашего партнера:
Полная версия книги
Всего 10 форматов

